Буферные системы крови норма

Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 28 августа 2018;
проверки требуют 15 правок.
Бу́ферные систе́мы кро́ви (от англ. buffer, buff — «смягчать удар») — физиологические системы и механизмы, обеспечивающие заданные параметры кислотно-основного равновесия в крови[1]. Они являются «первой линией защиты», препятствующей резким перепадам pH внутренней среды живых организмов.
Циркулирующая кровь представляет собой взвесь живых клеток в жидкой среде, химические свойства которой очень важны для их жизнедеятельности. У человека за норму принят диапазон колебаний pH крови 7,37—7,44 со средней величиной 7,4. Буферные системы крови слагаются из буферных систем плазмы и клеток крови и представлены следующими системами[1][2]:
- бикарбона́тная (водородкарбонатная) бу́ферная систе́ма;
- фосфа́тная бу́ферная систе́ма;
- белко́вая бу́ферная систе́ма;
- гемоглоби́новая бу́ферная система;
- эритроциты.
Помимо этих систем также активно участвуют дыхательная и мочевыделительная системы[1].
Бикарбонатная буферная система[править | править код]
Одна из самых мощных и вместе с тем самая управляемая система[2] внеклеточной жидкости и крови, на долю которой приходится около 53 % всей буферной ёмкости крови. Представляет собой сопряжённую кислотно-основную пару, состоящую из молекулы угольной кислоты H2CO3, являющейся источником протона, и бикарбонат-аниона HCO3−, выполняющего функцию акцептора протона:
Вследствие того, что концентрация гидрокарбоната натрия в крови значительно превышает концентрацию H2CO3, буферная ёмкость этой системы будет значительно выше по кислоте. Иначе говоря, гидрокарбонатная буферная система особенно эффективно компенсирует действие веществ, увеличивающих кислотность крови. К числу таких веществ прежде всего относят молочную кислоту, избыток которой образуется в результате интенсивной физической нагрузки. Гидрокарбонатная система наиболее «быстро» отзывается на изменение pH крови[2].
Фосфатная буферная система[править | править код]
В крови ёмкость фосфатной буферной системы невелика (составляет около 2 % общей буферной ёмкости), в связи с низким содержанием фосфатов в крови. Фосфатный буфер выполняет значительную функцию в поддержании физиологических значений рН во внутриклеточных жидкостях и моче.
Буфер образован неорганическими фосфатами. Функцию кислоты в этой системе выполняет однозамещённый фосфат (NaH2PО4), а функцию сопряженного основания — двузамещённый фосфат (Na2HPО4). При рН 7,4 соотношение [НРО42-/Н2РО4-] равняется поскольку при температуре 25+273,15K pKa, ортоII=7,21[3], при этом средний заряд аниона ортофосфорной кислоты < q >=((-2)*3+(-1)*2)/5=-1,4 единиц заряда позитрона.
Буферные свойства системы при увеличении в крови содержания водородных ионов реализуются за счет их связывания с ионами НРО42- с образованием Н2РО4-:
а при избытке ионов ОН- — за счет связывания их с ионами Н2РО4-:
Фосфатная буферная система крови тесно взаимосвязана с бикарбонатной буферной системой.
Белковая буферная система[править | править код]
В сравнении с другими буферными системами имеет меньшее значение для поддержания кислотно-основного равновесия (7—10 % буферной ёмкости).
Белки́ плазмы крови благодаря наличию кислотно-основных групп в молекулах белков (белок—H+ — кислота, источник протонов и белок− — сопряжённое основание, акцептор протонов) образуют буферную систему, наиболее эффективную в диапазоне pH 7,2—7,4[1].
Основную часть белков плазмы крови (около 90 %) составляют альбумины и глобулины. Изоэлектрические точки этих белков (число катионных и анионных групп одинаково, заряд молекулы белка равен нулю) лежат в слабокислой среде при pH 4,9—6,3, поэтому в физиологических условиях при pH 7,4 белки находятся преимущественно в формах «белок-основание» и «белок-соль».
Буферная ёмкость, определяемая белками плазмы, зависит от концентрации белков, их вторичной и третичной структуры и числа свободных протон-акцепторных групп. Эта система может нейтрализовать как кислые, так и основные продукты. Однако вследствие преобладания формы «белок-основание» её буферная ёмкость значительно выше по кислоте.
Буферная ёмкость свободных аминокислот плазмы крови незначительна как по кислоте, так и по щелочи. При физиологическом значении pH их мощность мала. Практически только одна аминокислота — гистидин — обладает значительным буферным действием при значении pH, близком к плазме крови.[2]
Эритроциты[править | править код]
Во внутренней среде эритроцитов в норме поддерживается постоянное значение pH, равное 7,30. Здесь также действуют гидрокарбонатная и фосфатная буферные системы. Однако их мощность отличается от таковой в плазме крови. Кроме того, в эритроцитах белковая система гемоглобин-оксигемоглобин играет важную роль как в процессе дыхания (транспортная функция по переносу кислорода к тканям и органам и удалению из них метаболической CO2), так и в поддержании постоянства pH внутри эритроцитов, а в результате и в крови в целом. Эта буферная система в эритроцитах тесно связана с гидрокарбонатной системой.[2]
Гемоглобиновая буферная система[править | править код]
Буферная система крови (75 % буферной ёмкости). Играет важную роль как в процессе дыхания (транспортная функция по переносу кислорода к тканям и органам и удалению из них метаболической CO2), так и в поддержании постоянства pH внутри эритроцитов, а в результате и в крови в целом.[2]
См. также[править | править код]
- Буферный раствор
Примечания[править | править код]
- ↑ 1 2 3 4 Березов Т. Т., Коровкин Б. Ф. Биологическая химия: Учебник — 1990 г. — стр. 452—455.
- ↑ 1 2 3 4 5 6 Ершов. Общая химия.Биофизическая химия.Химия биогенных элементов. — Издание восьмое, стериотипное. — Москва: Высшая школа, 2010. — 559 с. — ISBN 978-5-06-006180-2.
- ↑ И.Т.Гороновский, Ю.П.Назаренко, Е.Ф.Некряч. Краткий справочник по химии. — Пятое издание, исправленное и дополненное. — Киев: Наукова Думка, 1987. — С. 348. — 828 с.
Литература[править | править код]
- Березов Т. Т., Коровкин Б. Ф. [www.xumuk.ru/biologhim/ Биологическая химия: Учебник] / Под. ред. акад. АМН СССР С. С. Дебова.— 2-е изд., перераб. и доп.— М.: Медицина,— 1990.— 528 с., С. 452—455. ISBN 5-225-01515-8.
- Ершов. Общая химия.Биофизическая химия.Химия биогенных элементов. — Издание восьмое,стереотипное. — Москва: Высшая школа, 2010. — 559 с. — ISBN 978-5-06-006180-2.
Некоторые внешние ссылки в этой статье ведут на сайты, занесённые в спам-лист. Эти сайты могут нарушать авторские права, быть признаны неавторитетными источниками или по другим причинам быть запрещены в Википедии. Редакторам следует заменить такие ссылки ссылками на соответствующие правилам сайты или библиографическими ссылками на печатные источники либо удалить их (возможно, вместе с подтверждаемым ими содержимым). |
Источник
Установлено,
что состоянию нормы соответствует определенный диапазон колебаний рН крови – от
7,37 до 7,44 со средней величиной 7,40 . Кровь представляет собой взвесь
клеток в жидкой среде, поэтому ее кислотно-основное равновесие поддерживается
совместным участием буферных систем плазмы и клеток крови. Важнейшими буферными
системами крови являются бикарбонатная, фосфатная, белковая и наиболее мощная
гемогло-биновая.
Бикарбонатная буферная система – мощная и, пожалуй, самая управляемая система внеклеточной
жидкости и крови. На долю бикарбонатного буфера приходится около 10% всей
буферной емкости крови. Бикарбонатная система
представляет собой сопряженную кислотно-основную пару, состоящую из молекулы
угольной кислоты Н2СО3, выполняющую роль донора протона,
и бикарбонат-иона НСО3–, выполняющего роль акцептора
протона:
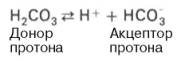
Для данной
буферной системы величину рН в растворе можно выразить через константу
диссоциации угольной кислоты (рКН2СО3) и логарифм концентрации
недиссоциированных молекул Н2СО3 и ионов HCO3–:
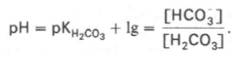
Истинная
концентрация недиссоциированных молекул Н2СО3 в крови
незначительна и находится в прямой зависимости от концентрации растворенного
углекислого газа (СО2 + Н2О <=> Н2СО3).
Поэтому удобнее пользоваться тем вариантом уравнения, в котором рКH2СО3
заменена «кажущейся» константой диссоциации Н2СО3,
учитывающей общую концентрацию растворенного СО2 в крови:
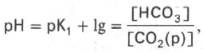
где K1– «кажущаяся» константа диссоциации Н2 С О3 ; [СО2(р)]
– концентрация растворенного СО2.
При
нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО3
в плазме крови превышает концентрацию СО2 примерно в 20 раз.
Бикарбонатная буферная система функционирует как эффективный регулятор в
области рН 7,4.
Механизм
действия данной системы заключается в том, что при выделении в кровь
относительно больших количеств кислых продуктов водородные ионы Н+
взаимодействуют с ионами бикарбоната НСО3–, что приводит
к образованию слабодиссоциирующей угольной кислоты Н2СО3.
Последующее снижение концентрации Н2СО3 достигается в
результате ускоренного выделения СО2 через легкие в результате их
гипервентиляции (напомним, что концентрация Н2СО3 в плазме
крови определяется давлением СО2 в альвеолярной газовой смеси).
Если в крови
увеличивается количество оснований, то они, взаимодействуя со слабой угольной
кислотой, образуют ионы бикарбоната и воду. При этом не происходит
сколько-нибудь заметных сдвигов в величине рН. Кроме того, для сохранения
нормального соотношения между компонентами буферной системы в этом случае
подключаются физиологические механизмы регуляции кислотно-основного равновесия:
происходит задержка в плазме
крови некоторого количества СО2 в результате гиповентиляции легких
. Как будет показано далее, данная буферная система тесно связана с
гемоглобиновой системой.
Фосфатная буферная система представляет собой сопряженную кислотно-основную пару,
состоящую из иона Н2РО4– (донор протонов) и
иона НРО42– (акцептор протонов):
Роль кислоты
в этой системе выполняет однозамещенный фосфат NaH2PO4,
а роль соли двузамещенный фосфат – Na2HPO4.
Фосфатная
буферная система составляет всего лишь 1% от буферной емкости крови. В других
тканях эта система является одной из основных. Для фосфатной буферной системы
справедливо следующее уравнение:
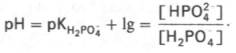
Во
внеклеточной жидкости, в том числе в крови, соотношение [НРО42–]:
[Н2РО4–] составляет 4:1. Величина рКН2РО4–
равна 6,86.
Буферное
действие фосфатной системы основано на возможности связывания водородных ионов
ионами НРО42– с образованием Н2РО4–
(Н+ + + НРО42– —> Н2РО4–),
а также ионов ОН– с ионами Н2РО4–
(ОН– + + Н2 Р О4– —> HPO42–+ H2O). Буферная пара (Н2РО4––НРО42–)
способна оказывать влияние при изменениях рН в интервале от 6,1 до 7,7 и может
обеспечивать определенную буферную емкость внутриклеточной жидкости, величина
рН которой в пределах 6,9–7,4. В крови максимальная емкость фосфатного буфера
проявляется вблизи значения рН 7,2. Фосфатный буфер в крови находится в тесном
взаимодействии с бикарбонатной буферной системой. Органические фосфаты также
обладают буферными свойствами, но мощность их слабее, чем неорганического
фосфатного буфера.
Белковая буферная система имеет меньшее значение для поддержания КОР в плазме крови,
чем другие буферные системы.
Белки
образуют буферную систему благодаря наличию кислотно-основных групп в молекуле
белков: белок–Н+ (кислота, донор протонов) и белок (сопряженное
основание, акцептор протонов). Белковая буферная система плазмы крови
эффективна в области значений рН 7,2–7,4.
Гемоглобиновая буферная система – самая мощная буферная система крови. Она в 9 раз мощнее
бикарбонатного буфера; на ее долю приходится 75% от всей буферной емкости
крови.
Участие
гемоглобина в регуляции рН крови связано с его ролью в транспорте кислорода и
углекислого газа. Константа диссоциации кислотных групп гемоглобина меняется в
зависимости от его насыщения кислородом. При насыщении кислородом гемоглобин
становится более сильной кислотой (ННbО2). Гемоглобин, отдавая
кислород, превращается в очень слабую органическую кислоту (ННb).
Итак,
гемоглобиновая буферная система состоит из неионизированного гемоглобина ННb
(слабая органическая кислота, донор протонов) и калиевой соли
гемоглобина КНb (сопряженное основание, акцептор протонов). Точно так же может
быть рассмотрена оксигемоглобиновая буферная система. Система гемоглобина и
система оксигемоглобина являются вза-имопревращающимися системами и существуют
как единое целое. Буферные свойства гемоглобина прежде всего обусловлены
возможностью взаимодействия кисло реагирующих соединений с калиевой солью
гемоглобина с образованием эквивалентного количества соответствующей калийной
соли кислоты и свободного гемоглобина:
КНb + Н2СO3—> КНСO3 + ННb.
Именно таким образом
превращение калийной соли гемоглобина эритроцитов в свободный ННb с
образованием эквивалентного количества бикарбоната обеспечивает поддержание рН
крови в пределах физиологически допустимых величин, несмотря на поступление в
венозную кровь огромного количества углекислого газа и других кисло реагирующих
продуктов обмена.
Гемоглобин
(ННb), попадая в капилляры легких, превращается в окси-гемоглобин (ННbО2),
что приводит к некоторому подкислению крови, вытеснению части Н2СО3
из бикарбонатов и понижению щелочного резерва крови . Перечисленные буферные
системы крови играют важную роль в регуляции кислотно-основного равновесия. Как
отмечалось, в этом процессе, помимо буферных систем крови, активное участие
принимают также система дыхания и мочевыделительная система.
Источник
Кровь играет определяющую роль в поддержании кислотно-щелочного равновесия, изменение которого может привести к развитию патологических состояний или гибели организма. Поэтому в организме существуют специальные системы, которые препятствуют изменению рН крови и других биологических жидкостей при образовании кислых и щелочных продуктов или при большом поступлении воды. Такую роль выполняют отдельные физиологические системы (дыхательная, выделительная), а также буферные системы. Последние очень быстро (в течение нескольких секунд) реагируют на изменение концентрации Н+ и ОН- в водных средах и являются срочными регуляторами кислотно-основного состояния в тканях организма.
Буферные системы – это смесь слабой кислоты и её растворимой соли, двух солей или белков, которые способны препятствовать изменению рН водных сред. Действие буферных систем направлено на связывание избытка Н+ или ОН- в среде и поддержание постоянства рН среды. При действии буферной системы образуются слабодиссоциируемые вещества или вода. К основным буферным системам крови относятся бикарбонатная, белковая (гемоглобиновая) и фосфатная. Имеются также ацетатная и аммонийная буферные системы.
Бикарбонатная буферная система – мощная и самая управляемая система крови и внеклеточной жидкости. На её долю приходится около 10% всей буферной ёмкости крови. Бикарбонатная система представляет собой сопряжённую кислотно-основную пару, состоящую из молекулы угольной кислоты Н2СО3, выполняющую роль донора протона, и бикарбонат-иона НСО3-, выполняющего роль акцептора протона:
СО2 + Н2О ↔ Н2СО3 ↔ Н+ + НСО3-
Истинная концентрация недиссоциированных молекул Н2СО3 в крови незначительна и находится в прямой зависимости от концентрации растворённого СО2. При нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО3- в плазме крови превышает концентрацию СО2 примерно в 20 раз. Бикарбонатная буферная система функционирует как эффективный регулятор в области рН = 7,4. Механизм действия этой системы заключается в том, что при выделении в кровь относительно больших количеств кислых продуктов протоны Н+ взаимодействуют с ионами бикарбоната НСО3- , что приводит к образованию слабодиссоциируемой Н2СО3.
Последующее снижение концентрации Н2СО3 достигается в результате ускоренного выделения СО2 через лёгкие в результате их гипервентиляции. Если в крови увеличивается количество оснований, то они, взаимодействуя со слабой угольной кислотой, образуют ионы бикарбоната и воду. При этом не происходит сколько-нибудь заметных сдвигов в величине рН. Кроме того, для сохранения нормального соотношения между компонентами буферной системы в этом случае подключаются физиологические механизмы регуляции кислотно-основного равновесия: происходит задержка в плазме крови некоторого количества СО2 в результате гиповентиляции лёгких. Бикарбонатная система тесно связана с гемоглобиновой системой.
Фосфатная буферная система представляет собой сопряжённую кислотно-основную пару, состоящую из иона Н2РО4- (донор протонов, выполняет роль кислоты) и иона НРО42- (акцептор протонов, выполняет роль соли). Фосфатная буферная система составляет лишь 1% от буерной ёмкости крови. В других тканях эта система является одной из основных. Фосфатная буферная система способна оказывать влияние при изменениях рН в интервале от 6,1 до 7,7 и может обеспечивать определённую ёмкость внутриклеточной жидкости, величина рН которой в пределах 6,9-7,4. В крови максимальная ёмкость фосфатного буфера проявляется вблизи значения 7,2. Органические фосфаты также обладают буферными свойствами, но мощность их слабее, чем неорганического фосфатного буфера.
Белковая буферная система имеет меньшее значение для поддержания кислотно-основного равновесия в плазме крови, чем другие буферные системы. Белки образуют буферную систему благодаря наличию кислотно-основных групп в молекуле белков: белок-Н+ (кислота, донор протонов) и белок (сопряжённое основание, акцептор протонов). Белковая буферная система плазмы крови эффективна в области значений рН 7,2-7,4.
Гемоглобиновая буферная система – самая мощная буферная система крови, на её долю приходится 75% от всей буферной. Участие гемоглобина в регуляции рН крови связано с его ролью в транспорте кислорода и углекислого газа. При насыщении кислородом гемоглобин становится более сильной кислотой (ННbО2). Гемоглобин, отдавая кислород, превращается в очень слабую органическую кислоту (ННb).
Гемоглобиновая буферная система состоит из неионизированного гемоглобина ННb (слабая органическая кислота, донор протонов) и калиевой соли гемоглобина КНb (сопряжённое основание, акцептор протонов). Точно так же может быть рассмотрена оксигемоглобиновая буферная система. Система гемоглобина и система оксигемоглобина являются взаимопревращающимися системами и существуют как единое целое. Буферные свойства гемоглобина обусловлены возможностью взаимодействия кисло реагирующих соединений с калиевой солью гемоглобина:
КНb + Н2СО3 => КНСО3 + ННb.
Это обеспечивает поддержание рН крови в пределах физиологически допустимых величин, несмотря на поступление в венозную кровь большого количества СО2 и других продуктов обмена кислотного характера. Гемоглобин (ННb), попадая в капилляры лёгких, превращается в оксигемоглобин (ННbО2), что приводит к некоторому подкислению крови, вытеснению части Н2СО3 из бикарбонатов и понижению щелочного резерва крови.
Дыхательная функция крови. Важной функцией крови является её способность переносить кислород к тканям и СО2 от тканей к лёгким. Веществом, осуществляющим эту функцию, является гемоглобин. Гемоглобин способен поглощать О2 при сравнительно высоком содержании его в атмосферном воздухе и легко отдавать при понижении парциального давления О2:
Нb + О2 ↔ НbО.
Поэтому в легочных капиллярах происходит насыщение крови О2, в то время как в тканевых капиллярах, где парциальное давление его резко снижается, наблюдается обратный процесс – отдача кровью кислорода тканям.
Образующийся в тканях при окислительных процессах СО2 подлежит выведению из организма. Обеспечение такого газообмена осуществляется несколькими системами организма.
Наибольшее значение имеют внешнее, или легочное, дыхание, обеспечивающее направленную диффузию газов через альвеолокапиллярные перегородки в легких и обмен газов между наружным воздухом и кровью; дыхательная функция крови, зависимая от способности плазмы растворять и способности гемоглобина обратимо связывать кислород и углекислый газ; транспортная функция сердечно-сосудистой системы (кровотока), обеспечивающая перенос газов крови от легких к тканям и обратно; функция ферментных систем, обеспечивающая обмен газов между кровью и клетками тканей, т.е. тканевое дыхание.
Диффузия газов крови осуществляется через мембрану клеток по концентрационному градиенту. За счет этого процесса в альвеолах легких в конце вдоха происходит выравнивание парциальных давлений различных газов в альвеолярном воздухе и крови. Обмен с атмосферным воздухом в процессе последующих выдоха и вдоха вновь приводит к различиям концентрации газов в альвеолярном воздухе и в крови, в связи с чем происходит диффузия кислорода в кровь, а углекислого газа из крови.
Большая часть О2 и СО2 переносится в форме связи их с гемоглобином в виде молекул HbO2 и HbCO2. Максимальное количество кислорода, связываемое кровью при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. В норме ее величина колеблется в пределах 16,0–24,0 об.% и зависит от содержания в крови гемоглобина, 1 г которого может связать 1,34 мл кислорода (число Хюфнера).
Связывание кислорода гемоглобином является обратимым процессом, зависимым от напряжения кислорода в крови, а также от других факторов, в частности от рН крови.
СО2, образующийся в тканях, переходит в кровь кровеносных капилляров, затем диффундирует внутрь эритроцита, где под влиянием карбоангидразы превращается в угольную кислоту, которая диссоциирует на Н+ и НСО3-. НСО3- частично диффундируют в плазму крови, образуя бикарбонат натрия. Он при поступлении крови в легкие (как и ионы НСО3-, содержащиеся в эритроцитах) образует СО2, который диффундирует в альвеолы.
Около 80% всего количества СО2 переносится от тканей к легким в виде бикарбонатов, 10% – в виде свободно растворенной углекислоты и 10% – в виде карбоксигемоглобина. Карбоксигемоглобин диссоциирует в легочных капиллярах на гемоглобин и свободный СО2, который удаляется с выдыхаемым воздухом. Освобождению СО2 из связи с гемоглобином способствует превращение последнего в оксигемоглобин, который, обладая выраженными кислотными свойствами, способен переводить бикарбонаты в угольную кислоту, диссоциирующую с образованием молекул воды и СО2.
При недостаточном насыщении крови кислородом развивается гипоксемия, которая сопровождается развитием гипоксии, т.е. недостаточным снабжением тканей кислородом. Тяжелые формы гипоксемии могут вызвать полное прекращение доставки кислорода тканям, тогда развивается аноксия, в этих случаях наступает потеря сознания, которая может закончиться смертью.
Патология газообмена, связанная с нарушением транспорта газов между легкими и клетками организма, наблюдается при уменьшении газовой емкости крови вследствие недостатка или качественных изменений гемоглобина, проявляется в виде анемических гипоксий. При анемиях кислородная емкость крови уменьшается пропорционально снижению концентрации гемоглобина. Снижение концентрации гемоглобина при анемиях ограничивает и транспорт углекислоты от тканей к легким в форме карбоксигемоглобина.
Нарушение транспорта кислорода кровью возникает также при патологии гемоглобина, например при серповидно-клеточной анемии, при инактивации части молекул гемоглобина за счет превращения его в метгемоглобин, например, при отравлении нитратами (метгемоглобинемия), или в карбоксигемоглобин (отравление СО).
Нарушения газообмена вследствие уменьшения объемной скорости кровотоки в капиллярах возникают при сердечной недостаточности, сосудистой недостаточности (в т.ч. при коллапсе, шоке), локальные нарушения – при ангиоспазме и др. В условиях застоя крови концентрация восстановленного гемоглобина возрастает. При сердечной недостаточности этот феномен особенно выражен в капиллярах отдаленных от сердца участков тела, где кровоток наиболее замедлен, что клинически проявляется акроцианозом.
Первичное нарушение газообмена на уровне клеток наблюдается главным образом при воздействии ядов, блокирующих дыхательные ферменты. В результате клетки утрачивают способность утилизировать кислород, и развивается резкая тканевая гипоксия, приводящая к структурной дезорганизации субклеточных и клеточных элементов, вплоть до некроза. Нарушению клеточного дыхания может способствовать витаминная недостаточность, например дефицит витаминов В2, РР, являющихся коферментами дыхательных ферментов.
Источник