Газообмен в легких гемоглобин

Одним из самых сложных процессов, что происходят в организме человека, несомненно, является дыхание. И сложность эта не только в танце легких, благодаря которому человек получает кислород, но и в процессах, с помощью которых этот кислород проникает дальше, в ткани, где превращается в углекислый газ, что отправляется в обратное путешествие. О данных процессах и пойдет речь далее.
Итак, приступим. Человек делает вдох, иии… Далеко не весь кислород поступает в легкие, а затем и в кровь. Часть вдыхаемого воздуха остается в так называемом мертвом пространстве. Мертвое пространство, в свою очередь, делится на анатомическое (дыхательные пути), в котором остается около 30 % вдыхаемого воздуха, и функциональное (вентилируемые, но по каким-то причинам не перфузируемые альвеолы).
Ухудшение альвеолярного газообмена может происходить при неглубоком и частом дыхании (причиной может стать перелом ребер, паралич дыхательной мускулатуры различного генеза и др.), а также при увеличении мертвого пространства, вызванном разнообразными причинами (нарушение перфузии альвеол в результате воспалительных заболеваний легких, удаление доли или целого легкого и др.), при снижении скорости кровотока по альвеолярным капиллярам (ТЭЛА, инфаркт легкого), при наличии диффузионного барьера (отек легких) и в результате ослабления альвеолярной вентиляции при обтурации просвета бронха. Газообмен между легкими и кровью происходит путем диффузии в соответствии с законом Фика. В легочных капиллярах она происходит за счет разности парциальных давлений в альвеолах и эритроцитах.
В альвеолах парциальное давление кислорода значительно превышает таковое для углекислого газа и составляет примерно 13,3 кПа (100 мм рт. ст.) и 5,3 кПа (40 мм рт. ст.) соответственно. Альвеолы омываются приносимой легочными артериями венозной кровью, в которой соотношение парциальных давлений этих двух газов обратно пропорционально и составляет приблизительно 5,3 кПа (40 мм рт. ст.) для кислорода и 6,1 кПа (46 мм рт. ст.) для углекислого газа. В среднем разница парциальных давлений составляет около 8 кПа (60 мм рт. ст.) для кислорода и около 0,8 кПа для углекислого газа.
Как уже было сказано выше, кислород путем диффузии проникает в кровь легочных капилляров. Диффузионное расстояние для кислорода при этом составляет 1–2 мкм, то есть именно на такое расстояние он проникает внутрь капилляра. Обмен крови в легочном капилляре происходит примерно за 0,75 секунды, но этого времени хватает на то, чтобы парциальные давления в альвеолах и в крови пришли в равновесие.
Кровь, в которой показатели парциального давления для кислорода и углекислого газа примерно равны таковым в альвеолах, называется артериализированной. Однако за счет наличия в легких артериовенозных шунтов и притока венозной крови из бронхиальных вен такой она остается недолго. В результате парциальное давление кислорода в аорте составляет примерно 12,0 кПа (как уже было сказано выше, парциальное давление в артериализированной крови равно таковому в альвеолах и составляет 13,3 кПа), а давление углекислого газа меняется незначительно и не приводит к затруднению его диффузии из крови в альвеолы.
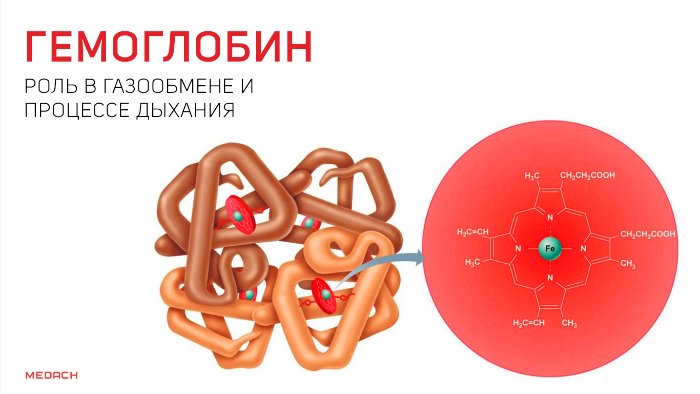
Но кислород непосредственно в ткани попадает лишь в крайне незначительных количествах: для свободного перемещения по организму ему необходим транспортер. Эту функцию выполняет содержащийся в эритроцитах белок — гемоглобин. Гемоглобин существует в оксигенированной и неоксигенированной формах. В дезокси-гемоглобине железо находится на уровне порфиринового кольца и стабилизируется электростатическими силами, что обеспечивает поддержание всей структуры. Появившись, кислород начинает «тянуть» за железо, которое переносится на проксимальный гистидин на другом конце полипептидной цепи и меняет структуру всего протеина.
В результате гемоглобин переходит в оксигенированную форму, альфа- и бета-цепи при этом поворачиваются относительно друг друга на 15 градусов, облегчая присоединение остальных молекул кислорода. В итоге каждый из четырех содержащихся в нем атомов двухвалентного железа обратимо связывается с молекулой кислорода, что превращает молекулу гемоглобина в оксигемоглобин. По сравнению с миоглобином гемоглобин имеет низкое сродство к кислороду, однако оно не статично. Так, миоглобин может связывать кислород только одним участком, поэтому кривая его связывания — гипербола. Кривая связывания гемоглобина с кислородом имеет S-образную форму, демонстрируя, что при его связывании с первой молекулой кислорода гемоглобин имеет очень низкое сродство к кислороду, но при связывании последующих молекул кислорода сродство остальных его субъединиц к нему значительно увеличивается и в конечном счете повышается примерно в 500 раз.
При этом альфа-цепи связывают кислород легче, чем бета-цепи. Этот процесс назван кооперативным взаимодействием. По мере снижения парциального давления кислорода в крови происходит его высвобождение из гемоглобина и поступление в ткани. Например, парциальное давление кислорода в работающих мышцах составляет всего 26 мм рт. ст, и при прохождении эритроцитов через капилляры, кровоснабжающие мышцы, происходит высвобождение и поступление в мышечные клетки примерно ⅓ всего переносимого гемоглобином кислорода. При повышении температуры тела также возрастает потребность в кислороде, что, в свою очередь, стимулирует высвобождение и поступление его в ткани. При снижении температуры, напротив, развивается гипоксия тканей, способствующая компенсаторному увеличению сродства гемоглобина к кислороду.
Гемоглобин также осуществляет перенос от тканей к легким продуктов тканевого дыхания — углекислого газа и ионов водорода. В ходе окислительных процессов в клетке выделяется углекислый газ, в результате гидратации которого образуются ионы водорода, что, в свою очередь, приводит к снижению рН. Давно известно, что снижение рН и повышение концентрации углекислого газа в крови оказывает сильное влияние на способность гемоглобина связывать кислород.
В периферических сосудах показатели рН низкие, и по мере связывания гемоглобина с ионами водорода и углекислым газом происходит снижение его сродства к кислороду. Это влияние величины рН и концентрации углекислого газа на способность гемоглобина связывать кислород называют эффектом Бора.
Обратная ситуация имеет место в альвеолярных капиллярах, где присоединение кислорода к гемоглобину превращает тот в более сильную кислоту.
При этом сродство гемоглобина к углекислому газу снижается, а повышение кислотности гемоглобина приводит к высвобождению излишка ионов водорода и образованию в крови из бикарбоната угольной кислоты, которая затем распадается на воду и углекислый газ. В обоих случаях углекислый газ из крови поступает в альвеолы, а затем в атмосферу. Данный процесс назван эффектом Холдейна. Стоит отметить, что важную роль в образовании углекислого газа в эритроцитах играет ион хлора, поступающий в плазму крови в обмен на бикарбонат при участии белка-переносчика АЕ1. Данный процесс в англоязычной литературе получил название «Chloride shift» или «перенос Хамбургера».
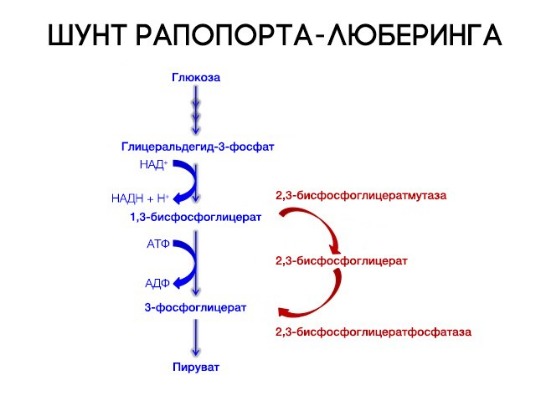
На сродство гемоглобина к кислороду оказывает влияние и присутствующее в эритроцитах вещество, получившее название 2,3-бисфосфоглицерат (БФГ). Его образование — своего рода побочная реакция анаэробного гликолиза, происходящего в эритроцитах в ходе ферментативного превращения глюкозы в пируват под действием фермента бифосфоглицератмутазы. БФГ способен самостоятельно связываться с неоксигенированной формой гемоглобина, образуя солевой мостик между двумя его бета-субъединицами и снижая сродство к кислороду.
При этом гемоглобин способен связать только одну молекулу БФГ, а при присоединении кислорода БФГ вытесняется из полости. В обычных условиях в эритроцитах крови содержится достаточно большое количество БФГ, которое может увеличиваться в условиях гипоксии (например, у дайверов при погружении на глубину), а также при восхождении на большую высоту. В первые часы подъема концентрация БФГ в эритроцитах будет возрастать, а сродство кислороду снижаться. Но на большой высоте парциальное давление будет значительно ниже такового на уровне моря, а значит, оно снизится и в тканях. При этом БФГ будет облегчать передачу кислорода от гемоглобина к тканям.
Некоторые вещества способны прочно связываться с гемоглобином или же вовсе менять его структуру. Одним из них является угарный газ, чье сродство к гемоглобину в 200 раз превышает таковое для кислорода. Отравления угарным газом часто происходят в помещениях с печным отоплением, при пожарах и авариях на производстве. Со временем кислород вытесняет угарный газ из гемоглобина, и в легких случаях пациенты помещаются под наблюдение и получают ингаляции с увлажненным кислородом. Необходимой мерой при тяжелых отравлениях угарным газом является переливание эритроцитарной массы.
К веществам, способным изменять структуру гемоглобина, относятся метгемоглобинобразователи — соединения, способные окислять двухвалентное железо в геме до трехвалентного. К ним относятся нитриты, нитраты, некоторые местные анестетики, аминофенолы, хлораты, примахин и некоторые сульфаниламиды. Состояние, характеризующееся появлением в крови окисленного гемоглобина, называют метгемоглобинемией. При высокой метгемоглобинемии капля крови, помещенная на фильтровальную бумагу, имеет характерный коричневый цвет, а при пропускании кислорода через пробирку с такой кровью ее цвет не меняется. Метгемоглобинемия выше 70 % от общего содержания гемоглобина часто приводит к гибели пациента еще до момента постановки диагноза.
Источники:
- Harrison’s hematology and oncology Longo, Dan L (Dan Louis), Third edition. New York : McGraw-Hill Education Medical, 2017.
- Наглядная физиология, С. Зильбернагль, А. Деспопулос, 2013.
- Ленинджер А. Основы биохимии: В 3-х т. Т. 1. /Д. Нельсон, М. Кокс ; Пер. с англ.-М.: БИНОМ: Лаборатория знаний, 2011.- 694 с.
Газообмен в легких
Биомеханика вдоха и выдоха
Вдох начинается с сокращения дыхательных (респираторных) мышц.
При вдохе межреберные мышцы сокращаются и приподнимают ребра, а диафрагма отодвигается в сторону брюшной полости, становится менее выпуклой. Сокращение диафрагмы приводит к увеличению объема грудной полости в вертикальном направлении, а сокращение наружных межреберных и межхрящевых мышц – к увеличению объема грудной полости в сагитальном и фронтальном направлениях. В результате этого объем грудной полости увеличивается. Так как давление в грудной полости ниже атмосферного, то при увеличении ее объема растягиваются и легкие. Давление в них на какой- то момент становится ниже атмосферного, в легкие по дыхательным путям устремляется воздух. При необходимости глубокого дыхания, кроме межреберных мышц и диафрагмы сокращаются, мышцы туловища и плечевого пояса.
Выдох осуществляется пассивно, в результате расслабления дыхательных мышц; он является следствием прекращения вдоха: межреберные мышцы расслабляются, ребра опускаются, диафрагма расслабляется, объем грудной полости и легких уменьшаются. Грудная клетка суживается под влиянием эластической тяги легких и постоянно имеющегося тонуса мышц стенки живота, при этом органы брюшной полости оказывают давление на диафрагму. Вследствие сужения грудной клетки легкие сжимаются. Давление в легких становится выше атмосферного: воздух выходит из них по дыхательным путям наружу.
Частота дыхания 16- 20 в минуту. Дыхательные движения грудной клетки обеспечивают вентиляцию альвеолярного воздуха и поддерживают постоянство его газового состава.
При глубоком вдохе происходит дополнительное сокращение межреберных и брюшных мышц и объем выдыхаемого воздуха возрастает. При глубоком, усиленном дыхании сокращаются не только главные мышцы, но и вспомогательные.
Объемы легочного воздуха
Человек в состоянии покоя вдыхает и выдыхает около 500 мл воздуха – это дыхательный объем. Если после спокойного вдоха сделать усиленный дополнительный вдох, то в легкие поступает еще 1500 мл воздуха – это резервный объем вдоха.
После спокойного выдоха можно при максимальном напряжении дыхательных мышц выдохнуть еще 1500 мл воздуха – это резервный объем выдоха.
В сумме дыхательный объем, резервный объем вдоха, и резервный объем выдоха составляют жизненную емкость легких.
ЖЕЛ = РОвд + ДО + РОвыд.
ЖЕЛ = 1500 + 500 + 1500 = 3500 мл.
ЖЕЛ – максимальный объем воздуха, который можно выдохнуть после глубокого вдоха.
После максимального выдоха в легких остается 1000 – 1500 мл воздуха – это остаточный объем. Его можно удалить не полностью только на трупе. В легочной ткани всегда остается воздух, поэтому, опущенный в воду кусочек легкого не тонет.
Функциональная остаточная емкость легких.
ФОЕЛ = РОвыд. + ОО
ФОЕЛ = 1500 + 1000 (1500) = 2500 – 3000 мл.
Емкость вдоха = РОвд. + ДО = 1500 + 500 = 2000 мл.
Объем воздуха, содержащийся в легких на высоте максимального вдоха, составляет Общую емкость легких.
ОЕЛ=ЖЕЛ + ОО
Воздух находится не только в альвеолах, но и в воздухоносных путях – полости носа, носоглотки, трахеи, бронхов. Воздух, находящийся в воздухоносных путях не участвует в газообмене, поэтому просвет воздухоносных путей называется мертвым пространством. Объем анатомического мертвого пространства около 150 мл.
Количественной характеристикой легочной вентиляции является минутный объем дыхания (МОД) – объем воздуха, проходящего через легкие за одну минуту. В состоянии покоя МОД равен 6- 9 л. При физической нагрузке его величина резко возрастает и составляет 25- 30 л.
Объем воздуха, который проходит через легкие за определенное время, называют максимальной вентиляцией легких (МВЛ). Этот параметр может достигать у молодого человека 120- 150 л/мин. МВЛ характеризует проходимость дыхательных путей, упругость грудной клетки и растяжимость легких.
Газообмен в легких
Газообмен осуществляется с помощью диффузии: СО2 выделяется из крови в альвеолы, О2 поступает из альвеол в венозную кровь, пришедшую в легочные капилляры из всех органов и тканей организма.

Рисунок 6. Газообмен между альвеолярным воздухом и эритроцитом. Цифрами обозначены величины парциального давления кислорода (PО2) и углекислого газа (РСО2) в артериальном и венозном конце капилляра
Процесс диффузии газов через альвеолярно-капиллярную мембрану зависит от следующих факторов:
1. градиента парциального давления газов по обе стороны мембраны (в альвеолярном воздухе и в легочных капиллярах);
2. толщины альвеолярно-капиллярной мембраны;
3. общей поверхности диффузии в легком (поверхность контакта легочных капилляров и альвеол составляет 60- 120 м2).
Парциальное давление кислорода в альвеолах (100 мм рт. ст.) значительно выше, чем напряжение, кислорода в венозной крови, поступающей в капилляры легких. Градиент парциального давления углекислого газа направлен в обратную сторону: 46 мм. рт. ст. в начале легочных капилляров и 40 мм рт. ст. в альвеолах. Эти градиенты давлений являются движущей силой диффузии кислорода и двуокиси углерода, т.е. газообмена в легких.
Согласно закону Фика диффузионный поток прямо пропорционален градиенту концентрации. Коэффициент диффузии для углекислого газа в 20- 25 раз больше, чем кислорода. При прочих равных условиях углекислый газ диффундирует через определенный слой среды в 20- 25 раз быстрее, чем кислород. Поэтому обмен СО2 в легких происходит достаточно полно, несмотря на небольшой градиент парциального давления этого газа.
При прохождении каждого эритроцита через легочные капилляры время, в течение которого возможна диффузия (время контакта) относительно невелико – около 0,3 сек. Однако, этого времени вполне достаточно для того, чтобы напряжения дыхательных газов в крови и их парциальное давление в альвеолах практически сравнялись.
Диффузионную способность легких, как и альвеолярную вентиляцию, следует рассматривать в отношении к перфузии (кровоснабжению) легких..
Транспорт газов кровью
Транспорт кислорода кровью.
Содержание растворенного газа в жидкости зависит от его парциального давления. Содержание в крови кислорода и углекислого газа в физически растворенном состоянии относительно невелико, однако это состояние играет существенную роль в жизнедеятельности организма.
Для того, чтобы связаться с теми или иными веществами, дыхательные газы сначала должны быть доставлены к ним в физически растворенном виде. Таким образом, при диффузии в ткани или кровь каждая молекула кислорода или углекислого газа определенное время пребывает в состоянии физического растворения. Большая часть кислорода переносится кровью в виде химического соединения с гемоглобином. 1 моль гемоглобина может связать до 4 молей кислорода, a 1 г гемоглобина – 1,39 мл кислорода. При анализе газового состава крови получают меньшую величину (1,34 – 1,36 мл О2 на 1 г Нb). Это обусловлено тем, что небольшая часть гемоглобина находится в неактивном виде. Таким образом, можно считать, что in vitro 1 г. Нb связывает 1,34 мл О2 – так называемое число Хюффнера.
Исходя из числа Хюффнера и зная содержание гемоглобина, вычислить кислородную емкость крови: 0,20 л кислорода на 1 л крови; Однако, такое содержание кислорода в крови может достигаться лишь в том случае, если кровь контактирует с газовой смесью с высоким содержанием кислорода (р О2 = 300 мм рт. ст.), поэтому в естественных условиях гемоглобин оксигенируется не полностью.
Реакция, отражающая соединение кислорода с гемоглобином, подчиняется закону действующих масс. Это означает, что отношение между количеством гемоглобина и оксигемоглобина зависит от содержания физически растворенного кислорода в крови; последнее же пропорционально напряжению кислорода. Процентное отношение оксигемоглобина к общему содержанию гемоглобина называется насыщением гемоглобина кислородом.
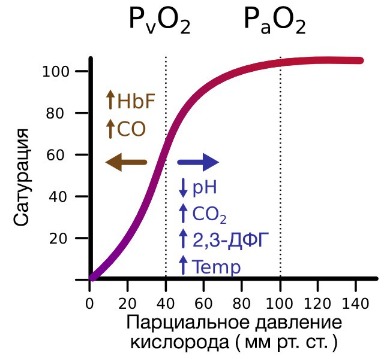
В соответствии с законом действующих масс насыщение гемоглобина кислородом зависит от напряжения кислорода. Графически эту зависимость отражает кривая диссоциации оксигемоглобина, которая имеет S- образную форму.

Рисунок 7. График диссоциации оксигемоглобина а – при нормальном парциальном давлении СО2 б – влияние изменений парциального давления СО2 в – влияние изменений pH; г – влияние изменений температуры.
Наиболее простым показателем, характеризующим расположение этой кривой служит так называемое напряжение полунасыщения рО2 т.е. такое напряжение кислорода, при котором насыщение гемоглобина кислородом составляет 50%. В норме рО2 артериальной крови составляет около 26 мм рт. ст.
Конфигурация кривой диссоциации оксигемоглобина имеет важное значение для переноса кислорода кровью. В процессе поглощения кислорода в легких напряжение О2 в крови приближается к парциальному давлению этого газа в альвеолах. У молодых людей рО2 артериальной крови составляет около 95 мм рт. ст. При таком напряжении насыщение гемоглобина кислородом равно примерно 97%. С возрастом и в ещё большей степени при заболеваниях легких напряжение кислорода в артериальной крови может значительно снижаться, однако, поскольку кривая диссоциации оксигемоглобина в правой части почти горизонтальна, насыщение крови кислородом уменьшается ненамного. Даже при падении напряжения кислорода в артериальной крови до 60 мм рт. ст., насыщение гемоглобина кислородом равно 90 %. Таким образом, благодаря тому, что области высоких напряжений кислорода соответствует горизонтальный участок кривой диссоциации оксигемоглобина, насыщение артериальной крови кислородом сохраняется на высоком уровне даже существенных сдвигах рО2.
Крутой наклон среднего участка кривой диссоциации оксигемоглобина свидетельствует о благоприятной ситуации для отдачи кислорода тканям. В состоянии покоя рО2 в области венозного конца капилляра равно приблизительно 40 мм рт. ст., что соответствует примерно 73% насыщения. Если в результате увеличения потребления кислорода его напряжение в венозной крови падает лишь на 5 мм рт. ст., то насыщение гемоглобина кислородом снижается на 7%; высвобождающийся при этом кислород может быть сразу же использован для процессов метаболизма.
Конфигурация кривой диссоциации оксигемоглобина обусловлена главным образом химическими свойствами гемоглобина; а также влиянием температуры, pH, напряжения углекислого газа. Как правило, эти факторы смещают кривую, увеличивая иди уменьшая ее наклон, но не изменяя при этом ее S – образную форму.
Если сродство гемоглобина к кислороду повышается, то процесс идет в сторону образования оксигемоглобина и график диссоциации смещается влево. При снижении сродства гемоглобина к кислороду процесс идет больше в сторону диссоциации оксигемоглобина, при этом график диссоциации смещается вправо. Равновесие реакции оксигенации гемоглобина зависит от температуры. При понижении температуры наклон кривой диссоциации оксигемоглобина увеличивается, а при ее повышении – снижается. У теплокровных животных этот эффект проявляется только яри гипотермии или лихорадочном состоянии.
Форма кривой диссоциации оксигемоглобина в значительной степени зависит от содержания в крови ионов Н+. При снижении pH (при закислении крови) сродство гемоглобина к кислороду уменьшается и кривая диссоциации оксигемоглобина уплощается. Влияние pH на расположение кривой диссоциации оксигемоглобина называется эффектом Бора.
Величина pH крови тесно связана с напряжением в ней СО2 (напряжение углекислого газа) – чем рСО2 выше, тем ниже pH. Увеличение напряжения в крови СО2 сопровождается снижением сродства гемоглобина к кислорода и уплощением кривой диссоциации оксигемоглобина. Эту зависимость также называют эффектом Бора, хотя при подробном количественном анализе было показано, что влияние углекислого газа на форму кривой нельзя объяснить только изменением pH. Очевидно, сам углекислый газ оказывает на диссоциацию оксигемоглобина «специфический эффект».
При ряде патологических состояний наблюдаются изменения процессов транспорта кислорода кровью. При некоторых видах анемий происходит сдвиг кривой вправо, реже влево. Известно, что на форму и расположение кривой оказывают выраженное влияние некоторые фосфорорганические соединения, содержание которых в эритроцитах при патологии может изменяться. Основным соединением является 2,3-дифосфоглицерат (2,3- ДФГ). Сродство гемоглобина к кислороду зависит от содержания в эритроцитах катионов. При патологических сдвигах pH также отмечаются соответствующие изменения: при алкалозе поглощение кислорода в легких в результате эффекта Бора увеличивается, но отдача его тканям затрудняется; при ацидозе наблюдается обратная картина. Значительный сдвиг кривой влево имеет место при отравлении угарным газом.
Транспорт углекислого газа кровью.
Двуокись углерода – конечный продукт окислительных обменных процессов в клетках – переносится с кровью к легким и удаляется через них во внешнюю среду. Так же как и кислород, СО2 может переноситься как в физически растворенном виде, так и в состоянии химических соединений.
Химические реакции связывания углекислого газа несколько сложнее, чем реакции присоединения кислорода. Это обусловлено тем, что механизмы, отвечающие за транспорт СО2, должны одновременно обеспечивать поддержание постоянства кислородно-основного равновесия крови и тем самым внутренней среды организма в целом.
Напряжение СО2 в артериальной крови, поступающей в тканевые капилляры составляет 40 мм рт. ст. В клетках же, расположенных около этих капилляров. Напряжение углекислого газа значительно выше, так как это вещество постоянно образуется в результате метаболизма. В связи с этим физически растворенный углекислый газ переносится по градиенту напряжения из тканей в капилляры. Здесь его некоторое количество остается в состоянии физического растворения. Но большая часть СО2 претерпевает, ряд химических превращений. Прежде всего, происходит гидратация молекул углекислого газа с образованием угольной кислоты.
В плазме крови эта реакция протекает очень медленно; в эритроците же она ускоряется примерно в 10000 раз, что связано с действием фермента карбоангидразы. Поскольку этот фермент присутствует только в клетках, практически все молекулы СО2, участвующие в реакции гидратации, должны сначала поступить в эритроциты. Следующая реакция в цепи химических превращений СО2 заключается в диссоциации слабой кислоты Н2СО2 на ионы бикарбоната и водорода.
Накопление НСО3– в эритроците приводит к тому, что между его внутренней средой и плазмой крови создается диффузионный градиент. Ионы НСО3– могут передвигаться по этому градиенту лишь в том случае, если при этом не будет нарушаться равновесное распределение электрических зарядов. В связи с этим одновременно с выходом каждого иона НСО3– должен происходить либо выход из эритроцита одного катиона, либо вход одного аниона.
Поскольку мембрана эритроцита практически непроницаема для катионов, но сравнительно легко пропускает небольшие анионы, взамен НСО3– в эритроцит поступают ионы хлора. Этот обменный процесс называется хлоридным сдвигом.
Углекислый газ может связываться также путем непосредственного присоединения к аминогруппам белкового компонента гемоглобина. При этом образуется так называемая карбаминовая связь. Гемоглобин, связанный с СО2, называется карбогемоглобином.

Рисунок 8. Схема процессов, происходящих в эритроцитах при поглощении или отдаче кровью кислорода и углекислого газа.
Содержание углекислого газа, находящегося в крови в виде химических соединений, непосредственно зависит от его напряжения. В свою очередь, величина рСО2 определяется скоростью образования СО2 в тканях и его выделения легкими. Зависимость содержания СО2 от его напряжения описывается кривой, аналогичной кривой диссоциации оксигемоглобина.
В легких происходит диссоциация соединений углекислого газа и выделение из организма углекислого газа. Начинается выход в альвеолы физически растворенного СО2 из плазмы крови, вследствие наличия градиента парциального давления РСО2 между альвеолами (40 мм.рт.ст.) и венозной кровью (46 мм.рт.ст.). Это ведет к уменьшению напряжения РСО2 в крови. Присоединение кислорода к гемоглобину ведет к уменьшению сродства углекислого газа к гемоглобину и расщепления карбогемогдобина.
Зависимость содержания СО2 от степени оксигенации гемоглобина называется эффектом Холдейна. Данный эффект частично обусловлен различной способностью оксигемоглобина и дезоксигемоглобина к образованию карбаминовой связи.
Газообмен а тканях
Газообмен в тканях так же, как и газообмен в легких зависит, от следующих факторов:
1. градиента напряжения газов между кровью и клетками
2. состояния мембран;
3. площади диффузии;
4. коэффициента диффузии.
В легких кровь из венозной превращается в артериальную:
– богатую кислородом
– бедную углекислым газом.
Артериальная кровь направляется к тканям, где в результате постоянно протекающих окислительных процессов потребляется кислород и образуется углекислый газ. В тканях напряжение кислорода близко к 0, а напряжение углекислого газа = 60 мм рт. ст.
Вследствие разности давления углекислый газ из ткани диффундирует в кровь, а кислород – в ткани. Кровь становится венозной, по венам поступает в легкие, где повторяется цикл обмена газов.