Гемоглобин образуется в результате обмена жиров

За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов.
Эритроциты в норме живут 90-120 дней, после чего лизируются в клетках ретикулоэндотелиальной системы – макрофагах селезенки (главным образом), купферовских клетках печени и макрофагах костного мозга. При разрушении эритроцитов в кровеносном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и также переносится в клетки РЭС селезенки, печени и костного мозга.
Синтез билирубина
В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина. Высвобождаемое железо может либо запасаться в клетке в комплексе с ферритином, либо выделяться наружу и связываться с трансферрином.
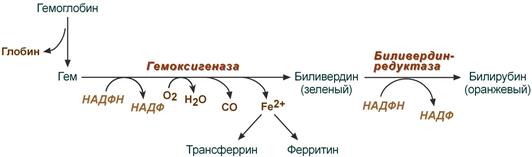
Реакции распада гемоглобина и образования билирубина
Билирубин – токсичное, жирорастворимое вещество, способное разобщать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
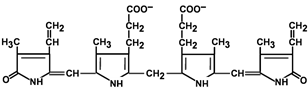
Строение билирубина
Выведение билирубина
Из клеток ретикуло-эндотелиальной системы билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.
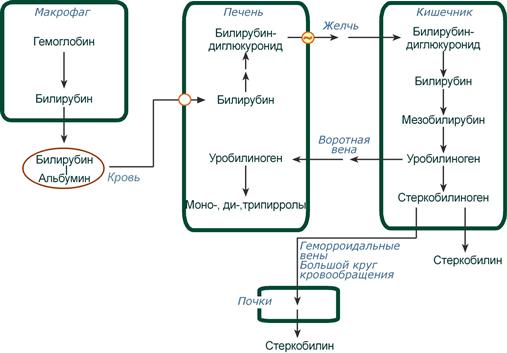
Этапы метаболизма билирубина в организме
Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (транспортный белок органических анионов) или по механизму флип-флоп. Далее при участии цитозольного связывающего белка лигандина (Y-протеин) билирубин транспортируется в ЭПР, где протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию конъюгации могут вступать сульфаты, фосфаты, глюкозиды.
Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин. Одновременно, даже в норме (особенно у взрослых), некоторое количество билирубин-глюкуронидов может попадать из желчи в кровь по межклеточным щелям.
Таким образом, в плазме крови обычно присутствуют две формы билирубина: свободный (непрямой), попадающий сюда из клеток РЭС (80% и более всего количества), и связанный (прямой), попадающий из желчных протоков (в норме не более 20%).
Термины “связанный“, “конъюгированный“, “свободный“, “несвязанный” отражают взаимодействие билирубина и глюкуроновой кислоты (но не билирубина и альбумина!).
Термины “прямой” и “непрямой” введены, исходя из возможности химической реакции билирубина с диазореактивом Эрлиха. Связанный билирубин реагирует с реактивом напрямую, без добавления дополнительных реагентов, т.к. является водорастворимым. Несвязанный (жирорастворимый) билирубин требует добавочных реактивов, реагирует не прямо.
Превращение в кишечнике
В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть уробилиногена всасывается и с кровью портальной вены попадает в печень, где либо распадается до моно-, ди- и трипирролов, либо окисляется до билирубина и снова экскретируется. При этом при здоровой печени в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена. Далее
- малая часть стеркобилиногена может всасываться и катаболизировать в печени, подобно уробилиногену,
- незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и в мочу. После окисления на воздухе из стеркобилиногена образуется стеркобилин мочи,
- однако основное количество стеркобилиногена достигает нижних отделов толстого кишечника и выделяется. В прямой кишке и на воздухе стеркобилиноген окисляется в стеркобилин, окрашивая кал,
- аналогично уробилиноген, появляющийся в моче при патологии печени, окисляется в уробилин.
Очень часто стеркобилиноген, содержащийся в нормальной моче, называют уробилиногеном. И в клинической практике обычно не проводят различий между стеркобилиногеном и уробилиногеном мочи, их рассматривают как один пигмент – урохромы (уробилиноиды), что может создавать некоторую путаницу при оценке результатов анализа.
Источник
Гемоглобин входит в состав группы белков гемопротеины, которые сами являются подвидом хромопротеинов и подразделяются на неферментативные белки (гемоглобин, миоглобин) и ферменты (цитохромы, каталаза, пероксидаза). Небелковой частью их является гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
Строение гемоглобина
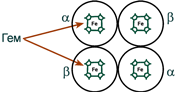
Строение гемоглобина А
Гемоглобин представляет собой белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями, при этом они взаимодействуют не произвольно, а определенным участком – контактной поверхностью. Этот процесс высокоспецифичен, контакт происходит одновременно в десятках точек по принципу комплементарности. Взаимодействие осуществляют разноименно заряженные группы, гидрофобные участки, неровности на поверхности белка.
Белковые субъединицы в нормальном гемоглобине могут быть представлены различными типами полипептидных цепей: α, β, γ, δ, ε, ξ (соответственно, греч. – альфа, бета, гамма, дельта, эпсилон, кси). В состав молекулы гемоглобина входят по две цепи двух разных типов.
Гем соединяется с белковой субъединицей, во-первых, через остаток гистидина координационной связью железа, во-вторых, через гидрофобные связи пиррольных колец и гидрофобных аминокислот. Гем располагается как бы “в кармане” своей цепи и формируется гемсодержащий протомер.
Нормальные формы гемоглобина
Существует несколько нормальных вариантов гемоглобина:
- HbР (primitive) – примитивный гемоглобин, содержит 2ξ- и 2ε-цепи, встречается в эмбрионе между 7-12 неделями жизни,
- HbF (foetal) – фетальный гемоглобин, содержит 2α- и 2γ-цепи, появляется через 12 недель внутриутробного развития и является основным после 3 месяцев,
- HbA (adult) – гемоглобин взрослых, доля составляет 98%, содержит 2α- и 2β-цепи, у плода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина,
- HbA2 – гемоглобин взрослых, доля составляет 2%, содержит 2α- и 2δ-цепи,
- HbO2 – оксигемоглобин, образуется при связывании кислорода в легких, в легочных венах его 94-98% от всего количества гемоглобина,
- HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
Патологические формы гемоглобина
HbS – гемоглобин серповидно-клеточной анемии.
MetHb – метгемоглобин, форма гемоглобина, включающая трехвалентный ион железа вместо двухвалентного. Такая форма образуется спонтанно, при взаимодействии молекулы O2 и гемового Fe2+, но обычно ферментативных мощностей клетки хватает на его восстановление. При использовании сульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления Fe3+ в Fe2+ в клинике используют аскорбиновую кислоту и метиленовую синь.
Hb-CO – карбоксигемоглобин, образуется при наличии СО (угарный газ) во вдыхаемом воздухе. Он постоянно присутствует в крови в малых концентрациях, но его доля может колебаться от условий и образа жизни.
Угарный газ является активным ингибитором гем-содержащих ферментов, в частности, цитохромоксидазы, 4-го комплекса дыхательной цепи.
Карбоксигемоглобин присутствует и в норме в количестве 0,5-1,5%, в сельской местности меньше, чем в городе. У курильщиков концентрация Hb-CO возрастает, в зависимости от количества сигарет в день, до 8-9%.
HbA1С – гликозилированный гемоглобин. Концентрация его нарастает при хронической гипергликемии и является хорошим скрининговым показателем уровня глюкозы крови за длительный период времени (время жизни эритроцита, 3-4 месяца).
Источник
РАЗДЕЛ 13. МЕТАБОЛИЗМ ГЕМА И ОБМЕН ЖЕЛЕЗА
III. Катаболизм гемоглобина
Эритроциты имеют короткое время жизни (примерно 120 дней). При физиологических условиях в организме взрослого человека разрушается около 1 — 2 x 1011 эритроцитов в сутки. Их катаболизм происходит главным образом в ретикулоэндотелиальных клетках селезёнки, лимфатических; узлов, костного мозга и печени. При старении эритроцитов снижается содержание сиаловых кислот в составе гликопротеинов плазматической мембраны. Изменённые углеводные компоненты гликопротеинов мембран эритроцитов связываются рецепторами клеток РЭС, и эритроциты «погружаются» в них эндоцитозом. Распад эритроцитов в этих клетках начинается с распада гемоглобина на гем и глобин и последующего гидролиза ферментами лизосом белковой части гемоглобина.
А. Катаболизм гема
Первая реакция катаболизма гема происходит при участии NADPH-зaвиcимoгo ферментативного комплекса гемоксигеназы. Ферментная система локализована в мембране ЭР, в области электронтранспортных цепей микросомального окисления. Фермент катализирует расщепление связи между двумя пиррольными кольцами, содержащих винильные остатки, — таким образом, раскрывается структура кольца (рис. 13-11). В ходе реакции образуются линейный тетрапиррол — биливердин (пигмент жёлтого цвета) и монооксид углерода (СО), который получается из углерода метениловой группы. Гем индуцирует транскрипцию гена гемоксигеназы, абсолютно специфичной по отношению к гему.
Рис. 13-11. Распад гема. М — (-СН3) — метильная группа; В — (-СН = СН2) — винильная группа; П — (-СН2-СН2-СООН) — остаток пропионовой кислоты. В ходе реакции одна метильная группа превращается в окись углерода и, таким образом, раскрывается структура кольца. Образованный биливердин под действием биливердинредуктазы превращается в билирубин.
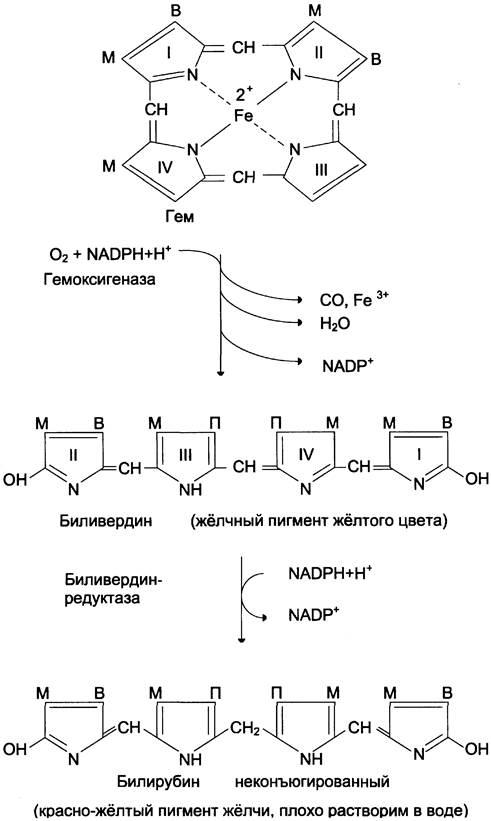
Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина NADPH-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, но также при катаболизме других гемсодержащих белков, таких как цитохромы и миоглобин. При распаде 1 г гемоглобина образуется 35 мг билирубина, а в сутки у взрослого человека — примерно 250 — 350 мг билирубина. Дальнейший метаболизм билирубина происходит в печени.
Б. Метаболизм билирубина
Билирубин, образованный в клетках РЭС (селезёнки и костного мозга), плохо растворим в воде, по крови транспортируется в комплексе с белком плазмы крови альбумином. Эту форму билирубина называют неконъюгированным билирубином. Каждая молекула альбумина связывает 2 (или даже 3) молекулы билирубина, одна из которых связана с белком более прочно (более высокое сродство), чем другие. При сдвиге pH крови в кислую сторону (повышение концентрации кетоновых тел, лактата) изменяются заряд, конформация альбумина, снижается сродство к билирубину. Поэтому билирубин, связанный с альбумином непрочно, может вытесняться из центров связывания и образовывать комплексы с коллагеном межклеточного матрикса и липидами мембран. Ряд лекарственных соединений конкурирует с билирубином за высокоаффинный, имеющий высокое сродство центр альбумина.
Поглощение билирубина паренхиматозными клетками печени
Комплекс «альбумин — билирубин», доставляемый с током крови в печень, на поверхности плазматической мембраны гепатоцита диссоциирует. Высвобожденный билирубин образует временный комплекс с липидами плазматической мембраны. Облегчённая диффузия билирубина в гепатоциты осуществляется двумя типами белков-переносчиков: лигандина (он транспортирует основное количество билирубина) и протеина Z. Активность поглощения билирубина гепатоцитом зависит от скорости его метаболизма в клетке.
Лигандин и протеин Z обнаружены также в клетках почек и кишечника, поэтому при недостаточности функции печени они способны компенсировать ослабление процессов детоксикации в этом органе.
Конъюгация билирубина в гладком ЭР
В гладком ЭР гепатоцитов к билирубину присоединяются (реакция конъюгации) полярные группы, главным образом от глюкуроновой кислоты. Билирубин имеет 2 карбоксильные группы, поэтому может соединяться с 2 молекулами глюкуроновой кислоты, образуя хорошо растворимый в воде конъюгат — диглюкуронид билирубина (конъюгированный, или прямой, билирубин) (рис. 13-12).
Рис. 13-12. Структура билирубиндиглюкуронида (конъюгированный, «прямой» билирубин). Глюкуроновая кислота присоединяется эфирной связью к двум остаткам пропионовой кислоты с образованием ацилглюкуронида.
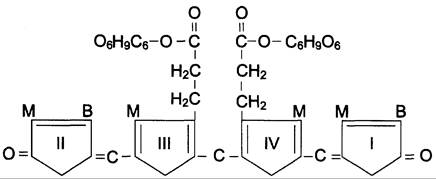
Донором глюкуроновой кислоты служит УДФ- глюкуронат. Специфические ферменты, УДФ-глюкуронилтрансферазы (уридиндифосфоглюкуронилтрансферазы) катализируют образование моно- и диглюкуронидов билирубина (рис. 13-13). Индукторами синтеза УДФ-глюкуронилтрансфераз служат некоторые лекарственные препараты, например, фенобарбитал (см. раздел 12).
Рис. 13-13. Образование билирубиндиглюкуронида.
Секреция билирубина в жёлчь
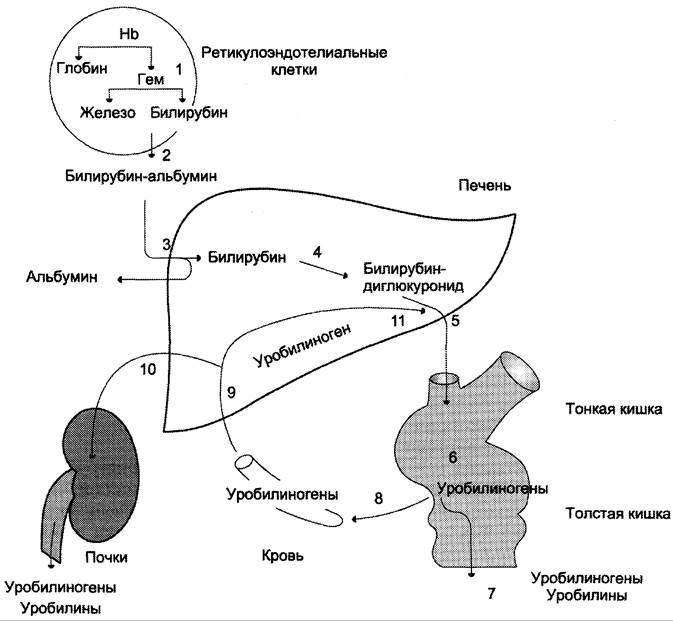
Секреция конъюгированного билирубина в жёлчь идёт по механизму активного транспорта, т. е. против градиента концентрации. Активный транспорт является, вероятно, скорость-лимитирующей стадией всего процесса метаболизма билирубина в печени. В норме диглюкуронид билирубина — главная форма экскреции билирубина в жёлчь, однако не исключается присутствие небольшого количества моноглюкуронида. Транспорт конъюгированного билирубина из печени в жёлчь активируется теми же лекарствами, которые способны индуцировать конъюгацию билирубина. Таким образом, можно сказать, что скорость конъюгации билирубина и активный транспорт би- лирубинглюкуронида из гепатоцитов в жёлчь строго взаимосвязаны (рис. 13-14).
Рис. 13-14. Билирубин-уробилиногеновый цикл в печени. 1 — катаболизм Нb в ретикулоэндотелиальных клетках костного мозга, селезёнки, лимфатических узлов; 2 — образование транспортной формы комплекса билирубин-альбумин; 3 — поступление билирубина в печень; 4 — образование билирубинглюкуронидов; 5 — секреция билирубина в составе жёлчи в кишечник; 6 — катаболизм билирубина под действием кишечных бактерий; 7 — удаление уробилиногенов с фекалиями; 8 — всасывание уробилиногенов в кровь; 9 — усвоение уробилиногенов печенью; 10 — поступление части уробилиногенов в кровь и выделение почками с мочой; 11 — небольшая часть уробилиногенов секретируется в жёлчь.
В. Катаболизм билирубиндиглюкуронида
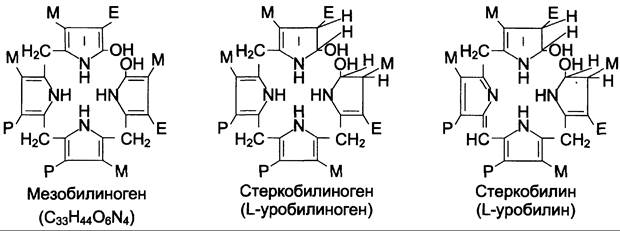
В кишечнике поступившие билирубинглюкурониды гидролизуются специфическими бактериальными ферментами р-глюкуронидазами, которые гидролизуют связь между билирубином и остатком глюкуроновой кислоты. Освободившийся в ходе этой реакции билирубин под действием кишечной микрофлоры восстанавливается с образованием группы бесцветных тетрапиррольных соединений — уробилиногенов (рис. 13-15).
Рис. 13-15. Структура некоторых жёлчных пигментов. Мезобилиноген — промежуточный продукт катаболизма билирубина в кишечнике.
В подвздошной и толстой кишках небольшая часть уробилиногенов снова всасывается, попадает с кровью воротной вены в печень. Основная часть уробилиногена из печени в составе жёлчи выводится в кишечник и выделяется с фекалиями из организма, часть уробилиногена из печени поступает в кровь и удаляется с мочой в форме уробилина (рис. 13-14). В норме большая часть бесцветных уробилиногенов, образующихся в толстой кишке, под действием кишечной микрофлоры окисляется в прямой кишке до пигмента коричневого цвета уробилина и удаляется с фекалиями. Цвет фекалий обусловлен присутствием уробилина.
Источник
Гемоглобин
освобождается (около 8-9г
в
день)
при
разрушении эритроцитов (время
их жизни
около
4
мес.). Оба
компонента гемоглобина (белок
и
гем) превращаются
далее независимо
друг от
друга.
Глобин протолитически гидролизуется
до
аминокислот,
а
гем
превращается в
пигменты жел-
чи.
Другие гемсодержащие белки (миоглобин,
ци-
тохромы,
пероксидазы, каталазы и
т.
д.)
разру-
шаются
аналогичным образом.
Гемоглобин,
освобождаемый из
эритроцитов,
сразу
же связывается
с
гаптоглобином, белком
плазмы
крови
(α2 – глобулин), одна
молекула кото-
рого
может связывать
две
молекулы Нb.
Образую-
щийся
комплекс адсорбируется ретикулоэндоте-
лиальной
системой
из
циркулирующей в
печени
и
других
органах крови.
В
комплексе
гаптоглобин—гемоглобин железо
гема
окисляется
до
Fe3+
(с
образованием
метгемоглобина),
окисление
α2
– метинового мостика
гема
до
формила
происходит
после
того, как
отщепляется
гаптоглобин.
При
открытии порфиринового
цикла
сродство
Fe3+
и
глобина
к
тетрапиррольному производному
понижается
и
молекула распадается
на линейное
производное
тетрапиррола биливердин,а
также
глобин
и
железо.
Биливердин
восстанавливается
NADPH по
цен
тральному
метиновому
мостику,
давая билирубин.
Билирубин
образует
гликозидную связь
с
UDP-
глюкуроновой
кислотой через
два
остатка
пропио-
новои
кислоты
центральных пиррольных
колец.
Этот
процесс
протекает в
эндоплазматическом ре-
тикулуме
и
катализируется
UDP-билирубин-глюку-
ронидтрансферазой.
Образуется
водорастворимый
глюкуронид
билирубина.
Будучи объемистым,
он
не
может проходить
обратно
через мембрану
клет-
ки
печени. Часть
билирубина
связывается с
серной
кислотой
под
действием
PAPS, а
также
с
глюкозой
или
ксилозой.
Таким
образом, его
молекула
стано-
вится
водорастворимой, и
в
этой
форме она
пере-
носится
в
желчные
протоки печени.
Вместе
с
желчью
она
попадает в
кишечник,
где превра-
щается
далее.
При
нарушении обмена тирозина может
развиться:
Тирозиноз
Алкаптонурия
Альбинизм
Микседема
Фенилкетонурия
Фенилкетонурия
характеризуется избыточным образованием:
Гомогентизиновой
кислотыМеланина
Фенилпировиноградной
кислотыМалеилацетоуксусной
кислотыФумаровой
кислоты
Причинами
положительного азотистого баланса
являются:
Рост
Беременность
Лихорадка
Ожоги
Повышение
содержания глюкокортикоидов
Для
образования каких биологически
активных веществ необходим гистидин:
Тиреоидных
гормоновМеланина
Гистамина
Холина
Адреналина
Какие
из аминокислот являются медиаторами
тормозного типа:
Аспартат
Глутамат
Глицин
ГАМК
Дофамин
Какие
из аминокислот относятся к медиаторам
возбуждающего типа:
Аспартат
Глутамат
Глицин
ГАМК
Дофамин
Таурин
При
гиперпродукции какого гормона
происходит повышение основного обмена:
АКТГ
Инсулин
Паратгормон
Вазопрессин
Тироксин
Фенилкетонурия
является нарушением обмена:
Тирозина
Фенилаланина
Гомогентизиновой
кислоты
Фенилкетонурия
характеризуется избыточным образованием:
Гомогентизиновой
кислотыДОФА
Меланина
Фенилпировиноградной
кислотыМалеилацетоуксусной
кислотыФумаровой
кислоты
Причинами
положительного азотистого баланса
являются:
Рост
Беременность
Лихорадка
Повышение
содержания глюкокортикоидовБолезни
почек
Положительный
азотистый баланс наблюдается при
повышенном образовании:
Cоматотропного
гормонаИнсулина
Глюкокортикоидов
Тироксина
Половых
гормонов
Отрицательный
азотистый баланс наблюдается при
недостатке:
Половых
гормоновИнсулина
Тироксина
Глюкокортикоидов
Жировая
дистрофия печени развивается при
дефиците:
Гистидина
Глутамина
Метионина
Таурина
Для
образования каких биологически
активных веществ необходим гистидин:
Тиреоидных
гормоновМеланина
Гистамина
Холина
Адреналина
Какие
из аминокислот относятся к медиаторам
возбуждающего типа:
Аспартат
Глутамат
Глицин
ГАМК
Дофамин
Таурин
Первый
период голодания характеризуется
преимущественным расходованием:
Углеводов
Жиров
Белков
Воды
Витаминов
Второй
период голодания характеризуется
преимущественным расходованием:
Углеводов
Жиров
Белков
Третий
период голодания характеризуется
преимущественным распадом:
Углеводов
Жиров
Белков
Ниболее
интенсивно происходит снижение во
время голодания массы:
Жировой
тканиМышц
Костей
Нервной
тканиСердца
Наименее
интенсивно происходит снижение во
время голодания массы:
Селезенки
Печени
Жировой
тканиМышц
Нервной
тканиСердца
Какая
аминокислота является источником
оксида азота:
Аспарагиновая
Глутаминовая
L-Аргинин
Аланин
Конечным
продуктом распада белков являются:
Молочная
кислотаГлютаминовая
кислотаАммиак
Пировитноградная
кислота
Гиперазотемия
может быть:
Гиперлактацидемическая
Кетоацидотическая
Гипохлоремическая
гипергликемическая
Голодание
называют частичным, если в организм
не поступают
Только
белкиТолько
водаЛюбой
из питательных веществТолько
минеральные веществаПоступление
пищи ограничено
Какие
белковые фракции преобладают в плазме
крови
Фибриноген
Альфа-1-глобулины
Бетта
–глобулиныАльбумины
Альфа-2-глобулины
Гамма-глобулинны
К
парапротеинам относятся:
Альбумины
Гамма-глобулины
Криоглобулины
Альфа-глобулины
В
первый период голодания увеличивается:
Гликогенез
Синтез
белкаЛиполиз
Гликогенолиз
Третий
период голодания характеризуется:
Усилением распада
гликогенаКатаболизмом
белкаРаспадом
жировПерераспределением
белков
Какие
процессы приводят к образованию новых
аминокислот:
трансаминирование
дезаминирование
декарбоксилирование
Положительный
азотистый баланс наблюдается при:
Лихорадке
Гипертиреозе
Голодании
В
стадию выздоровления
В
желудке расщепление белков происходит
при участии:
Пепсина
Химотрипсина
Энтерокиназы
Дипептидазы
В
кишечнике расщепление белков происходит
при участии:
Пепсина
Химотрипсина
Реннина
Энтерокиназы
Дипептидазы
Целиакия
– это заболевание, связанное с
недостаточностью расщепления:
Белков
мясаЛипидов
Сахарозы
Белков
злаковКрахмала
К
реакция межуточного обмена белков
относятся реакции:
Переаминирования
Глюконеогенеза
Β-окисления
Декарбоксилирования
Кетообразования
В
результате дезаминирования образуется:
Аммиак
Мочевая
кислотаМочевина
Причинами
повышения дезаминирования являются:
Голодание
Сахарный
диабетИзбыточное
потребление аминокислотНедостаток
витамина В6
При
недостатке витамина B6
происходят нарушения реакций:
Дезаминирования
Декарбоксилирования
Трансаминирования
Последствиями
уменьшения реакций дезаминирования
являются:
Уменьшение
синтеза белкаСнижение
активности цикла КребсаПовышение
содержания аммиака
Синтез
заменимых аминокислот происходит в
реакциях:
Дезаминирования
Декарбоксилирования
Трансаминирования
Образование
биогенных аминов происходит в реакциях:
Дезаминирования
Декарбоксилирования
Трансаминирования
Остаточный
азот – это все азотсодержащие вещества
плазмы крови, кроме:
Белка
Пептидов
Пептидов
и аминокислотАминокислот
Главным
компонентом остаточного азота плазмы
крови является:
Аминокислоты
Аммиак
Ионы
аммонияМочевина
Гиперазотемии
бывают:
Ретенционные
Гипохлоремические
Гиперхлоремические
Гипоаммониемия
Гипераммониемия
Желудочные
Предрасполагающими
к возникновению подагры факторами
являются:
Мясная
пищаМужской
полЖенский
полУпотребление
кофеПовышенная
активность ксантиноксидазыЩелочное
питье
Ретенционная
гиперазотемия является следствием
Болезней
почекТромбоза
воротной веныШока
Лихорадки
Обезвоживания
Продукционная
гиперазотемия характеризуется
увеличением в плазме крови:
Аммиака
Белка
Аминокислот
Креатина
При
каком из видов гиперазотемий в крови
снижено содержание мочевины:
Ретенционная
Гипохлоремическая
Гипераммониемия
Печеночная
Подагра
– это заболевание, связанное с
нарушением обмена:
Протаминов
Гистонов
Нуклеопротеинов
Мочевины
Пуриновых
оснований
С
распадом каких азотистых оснований
связано возникновение подагры:
Гуанина
Урацила
Тимидина
Аденина
Урацила
Подагра
– это заболевание:
Костей
Почек
Сердца
Суставов
печени
Гипераммониемия
является результатом снижения
активности:
Орнитинкарбамаилтрансферазы
Аргининсукцинатсинтетазы
Аргининсукцинатлиазы
цитохромоксидазы
Отрицательный
азотистый баланс развивается при:
Ожирении
Исхудании
Сердечной
недостаточностиИзбытке
инсулинаКровопотере
Положительный
азотистый баланс наблюдается при:
Лихорадке
Голодании
В стадию
выздоровленияГипертиреозе
При
наследственных заболеваниях наиболее
часто нарушен обмен
Валина
Фенилаланина
Изолейцина
Лейцина
Конечным
продуктам распада белков являются
Молочная
кислотаГлютаминовая
кислотаАммиак
Пировиноградная
кислота
Гиперазотемия
может быть
Гиперлактацидемическая
Кетоацидотическая
Гипохлоремическая
Гипергликемическая
Какие
белковые фракции преобладают в плазме
крови?
Фибриноген
Альфа-1-глобулины
Бетта-глобулины
Альбумины
Дыхательный
коэффициент в первый период голодания:
Снижается
Резко
снижаетсяПовышается
Не изменяется
Источник