Гемоглобин строение характеристика белкового и небелкового компонентов
Молекула гемоглобина: 4 субъединицы окрашены в разные цвета
Структура гемоглобина человека. Железосодержащие гем-группы показаны зелёным. Красным и синим показаны альфа- и бета- субъединицы.
Гемоглоби́н (от др.-греч. αἷμα «кровь» + лат. globus «шар») (Hb или Hgb) — сложный железосодержащий белок животных, обладающих кровообращением, способный обратимо связываться с кислородом, обеспечивая его перенос в ткани. У позвоночных животных содержится в эритроцитах, у большинства беспозвоночных растворён в плазме крови (эритрокруорин) и может присутствовать в других тканях[1]. Молекулярная масса гемоглобина человека — около 66,8 кДа. Молекула гемоглобина может нести до четырёх молекул кислорода. Один грамм гемоглобина может переносить до 1.34 мл. O2
Гемоглобин появился более чем 400 миллионов лет назад у последнего общего предка человека и акул в результате 2 мутаций, приведших к формированию четырёхкомпонентного комплекса гемоглобина, сродство которого к кислороду достаточно для связывания кислорода в насыщенной им среде, но недостаточно, чтобы удерживать его в других тканях организма.[2][3]
Большой вклад в исследование структуры и функционирования гемоглобина внёс Макс Фердинанд Перуц, получивший за это в 1962 году Нобелевскую премию[4].
Нормальным содержанием гемоглобина в крови человека считается: у мужчин — 130—160 г/л (нижний предел — 120, верхний предел — 180 г/л), у женщин — 120—160 г/л; у детей нормальный уровень гемоглобина зависит от возраста и подвержен значительным колебаниям. Так, у детей через 1—3 дня после рождения нормальный уровень гемоглобина максимален и составляет 145—225 г/л, а к 3—6 месяцам снижается до минимального уровня — 95—135 г/л, затем с 1 года до 18 лет отмечается постепенное увеличение нормального уровня гемоглобина в крови[5].
Во время беременности в организме женщины происходит задержка и накопление жидкости, что является причиной гемодилюции — физиологического разведения крови. В результате наблюдается относительное снижение концентрации гемоглобина (при беременности уровень гемоглобина в норме составляет 110—155 г/л). Кроме этого, в связи с внутриутробным ростом ребёнка происходит быстрое расходование запасов железа и фолиевой кислоты. Если до беременности у женщины был дефицит этих веществ, проблемы, связанные со снижением гемоглобина, могут возникнуть уже на ранних сроках беременности[6].
Главные функции гемоглобина: перенос кислорода и буферная функция. У человека в капиллярах лёгких в условиях избытка кислорода последний соединяется с гемоглобином. Потоком крови эритроциты, содержащие молекулы гемоглобина со связанным кислородом, доставляются к органам и тканям, где кислорода мало; здесь необходимый для протекания окислительных процессов кислород освобождается от связи с гемоглобином. Кроме того, гемоглобин способен связывать в тканях небольшое количество диоксида углерода (CO2) и освобождать его в лёгких.
Монооксид углерода (CO) связывается с гемоглобином крови намного сильнее (в 250 раз[7]), чем кислород, образуя карбоксигемоглобин (HbCO). Впрочем, монооксид углерода может быть частично вытеснен из гема при повышении парциального давления кислорода в лёгких. Некоторые процессы приводят к окислению иона железа в гемоглобине до степени окисления +3. В результате образуется форма гемоглобина, известная как метгемоглобин (HbOH) (metHb, от «мета-» и «гемоглобин», иначе гемиглобин или ферригемоглобин, см. Метгемоглобинемия). В обоих случаях блокируются процессы транспортировки кислорода.
Строение[править | править код]
Гемоглобин является сложным белком класса гемопротеинов, то есть в качестве простетической группы здесь выступает гем — порфириновое ядро, содержащее железо. Гемоглобин человека является тетрамером, то есть состоит из 4 протомеров. У взрослого человека они представлены полипептидными цепями α1, α2, β1 и β2. Субъединицы соединены друг с другом по принципу изологического тетраэдра. Основной вклад во взаимодействие субъединиц вносят гидрофобные взаимодействия. И α-, и β-цепи относятся к α-спиральному структурному классу, так как содержат исключительно α-спирали. Каждая цепь содержит восемь спиральных участков, обозначаемых буквами от A до H (от N-конца к C-концу).
Гем представляет собой комплекс протопорфирина IX, относящегося к классу порфириновых соединений, с атомом железа(II). Этот кофактор нековалентно связан с гидрофобной впадиной молекул гемоглобина и миоглобина.
Железо(II) характеризуется октаэдрической координацией, то есть связывается с шестью лигандами. Четыре из них представлены атомами азота порфиринового кольца, лежащими в одной плоскости. Две другие координационные позиции лежат на оси, перпендикулярной плоскости порфирина. Одна из них занята азотом остатка гистидина в 93-м положении полипептидной цепи (участок F). Связываемая гемоглобином молекула кислорода координируется к железу с обратной стороны и оказывается заключённой между атомом железа и азотом ещё одного остатка гистидина, располагающегося в 64-м положении цепи (участок E).
Всего в гемоглобине человека четыре участка связывания кислорода (по одному гему на каждую субъединицу), то есть одновременно может связываться четыре молекулы. Гемоглобин в лёгких при высоком парциальном давлении кислорода соединяется с ним, образуя оксигемоглобин. При этом кислород соединяется с гемом, присоединяясь к железу гема на 6-ю координационную связь. На эту же связь присоединяется и монооксид углерода, вступая с кислородом в «конкурентную борьбу» за связь с гемоглобином, образуя карбоксигемоглобин.
Связь гемоглобина с монооксидом углерода более прочная, чем с кислородом. Поэтому часть гемоглобина, образующая комплекс с монооксидом углерода, не участвует в транспорте кислорода. В норме у человека образуется 1,2 % карбоксигемоглобина. Повышение его уровня характерно для гемолитических процессов, в связи с этим уровень карбоксигемоглобина является показателем гемолиза.
Физиология[править | править код]
Изменение состояний окси- и дезоксигемоглобина
В отличие от миоглобина гемоглобин имеет четвертичную структуру, которая придаёт ему способность регулировать присоединение и отщепление кислорода и характерную кооперативность: после присоединения первой молекулы кислорода связывание последующих облегчается. Структура может находиться в двух устойчивых состояниях (конформациях): оксигемоглобин (содержит 4 молекулы кислорода; напряжённая конформация) и дезоксигемоглобин (кислорода не содержит; расслабленная конформация).
Устойчивое состояние структуры дезоксигемоглобина усложняет присоединение к нему кислорода. Поэтому для начала реакции необходимо достаточное парциальное давление кислорода, что возможно в альвеолах лёгких. Изменения в одной из 4-х субъединиц влияет на оставшиеся, и после присоединения первой молекулы кислорода связывание последующих облегчается.
Отдав кислород тканям, гемоглобин присоединяет к себе ионы водорода и углекислый газ, перенося их в лёгкие[8].
Гемоглобин является одним из основных белков, которыми питаются малярийные плазмодии — возбудители малярии, и в эндемичных по малярии районах земного шара весьма распространены наследственные аномалии строения гемоглобина, затрудняющие малярийным плазмодиям питание этим белком и проникновение в эритроцит. В частности, к таким имеющим эволюционно-приспособительное значение мутациям относится аномалия гемоглобина, приводящая к серповидноклеточной анемии. Однако, к несчастью, эти аномалии (как и аномалии строения гемоглобина, не имеющие явно приспособительного значения) сопровождаются нарушением кислород-транспортирующей функции гемоглобина, снижением устойчивости эритроцитов к разрушению, анемией и другими негативными последствиями. Аномалии строения гемоглобина называются гемоглобинопатиями.
Гемоглобин высокотоксичен при попадании значительного его количества из эритроцитов в плазму крови (что происходит при массивном внутрисосудистом гемолизе, геморрагическом шоке, гемолитических анемиях, переливании несовместимой крови и других патологических состояниях). Токсичность гемоглобина, находящегося вне эритроцитов, в свободном состоянии в плазме крови, проявляется тканевой гипоксией — ухудшением кислородного снабжения тканей, перегрузкой организма продуктами разрушения гемоглобина — железом, билирубином, порфиринами с развитием желтухи или острой порфирии, закупоркой почечных канальцев крупными молекулами гемоглобина с развитием некроза почечных канальцев и острой почечной недостаточности.
Ввиду высокой токсичности свободного гемоглобина в организме существуют специальные системы для его связывания и обезвреживания. В частности, одним из компонентов системы обезвреживания гемоглобина является особый плазменный белок гаптоглобин, специфически связывающий свободный глобин и глобин в составе гемоглобина. Комплекс гаптоглобина и глобина (или гемоглобина) затем захватывается селезёнкой и макрофагами тканевой ретикуло-эндотелиальной системы и обезвреживается.
Другой частью гемоглобинообезвреживающей системы является белок гемопексин[en], специфически связывающий свободный гем и гем в составе гемоглобина. Комплекс гема (или гемоглобина) и гемопексина затем захватывается печенью, гем отщепляется и используется для синтеза билирубина и других жёлчных пигментов, или выпускается в рециркуляцию в комплексе с трансферринами для повторного использования костным мозгом в процессе эритропоэза.
Экспрессия генов гемоглобина до и после рождения.
Также указаны типы клеток и органы, в которых происходит экспрессия гена (данные по Wood W. G., (1976). Br. Med. Bull. 32, 282.).[9]
Гемоглобин при заболеваниях крови[править | править код]
Дефицит гемоглобина может быть вызван, во-первых, уменьшением количества молекул самого гемоглобина (см. анемия), во-вторых, из-за уменьшенной способности каждой молекулы связать кислород при том же самом парциальном давлении кислорода.
Гипоксемия — это уменьшение парциального давления кислорода в крови, её следует отличать от дефицита гемоглобина. Хотя и гипоксемия, и дефицит гемоглобина являются причинами гипоксии.
Если дефицит кислорода в организме в общем называют гипоксией, то местные нарушения кислородоснабжения называют ишемией.
Прочие причины низкого гемоглобина разнообразны: кровопотеря, пищевой дефицит, болезни костного мозга, химиотерапия, отказ почек, атипичный гемоглобин.
Повышенное содержание гемоглобина в крови связано с увеличением количества или размеров эритроцитов, что наблюдается также при истинной полицитемии. Это повышение может быть вызвано: врождённой болезнью сердца, лёгочным фиброзом, слишком большим количеством эритропоэтина.
См. также[править | править код]
- Гемоглобин А
- Гемоглобин С (мутантная форма)
- Эмбриональный Гемоглобин (эмбриональный)
- Гемоглобин S (мутантная форма)
- Гемоглобин F (фетальный)
- Кобоглобин
- Нейроглобин
- Анемия
- Порфирия
- Талассемия
- Эффект Вериго — Бора
Примечания[править | править код]
- ↑ Haemoglobins of invertebrate tissues. Nerve haemoglobins of Aphrodite, Aplysia and Halosydna
- ↑ Ученые выяснили происхождение гемоглобина. РИА Новостей, 20.05.2020, 18:59
- ↑ Michael Berenbrink. Evolution of a molecular machine/Nature, NEWS AND VIEWS, 20 MAY 2020
- ↑ Лауреаты нобелевской премии. Макс Перуц.
- ↑ Назаренко Г. И., Кишкун А. А. Клиническая оценка результатов лабораторных исследований. — 2005.
- ↑ Общий анализ крови и беременность Архивная копия от 10 марта 2014 на Wayback Machine
- ↑ Hall, John E. Guyton and Hall textbook of medical physiology (англ.). — 12th ed.. — Philadelphia, Pa.: Saunders/Elsevier, 2010. — P. 1120. — ISBN 978-1416045748.
- ↑ Степанов В. М. Структура и функции белков : Учебник. — М. : Высшая школа, 1996. — С. 167—175. — 335 с. — 5000 экз. — ISBN 5-06-002573-X.
- ↑ Айала Ф., . Современная генетика: В 3-х т = Modern Genetics / Пер. А. Г. Имашевой, А. Л. Остермана, . Под ред. Е. В. Ананьева. — М.: Мир, 1987. — Т. 2. — 368 с. — 15 000 экз. — ISBN 5-03-000495-5.
Литература[править | править код]
- Mathews, CK; KE van Holde & KG Ahern (2000), Biochemistry (3rd ed.), Addison Wesley Longman, ISBN 0-8053-3066-6
- Levitt, M & C Chothia (1976), “Structural patterns in globular proteins”, Nature
Ссылки[править | править код]
- Eshaghian, S; Horwich, TB; Fonarow, GC (2006). “An unexpected inverse relationship between HbA1c levels and mortality in patients with diabetes and advanced systolic heart failure”. Am Heart J. 151 (1): 91.e1—91.e6. DOI:10.1016/j.ahj.2005.10.008. PMID 16368297.
- Kneipp J, Balakrishnan G, Chen R, Shen TJ, Sahu SC, Ho NT, Giovannelli JL, Simplaceanu V, Ho C, Spiro T (2005). “Dynamics of allostery in hemoglobin: roles of the penultimate tyrosine H bonds”. J Mol Biol. 356 (2): 335—53. DOI:10.1016/j.jmb.2005.11.006. PMID 16368110.
- Hardison, Ross C. (2012). “Evolution of Hemoglobin and Its Genes”. Cold Spring Harbor Perspectives in Medicine. 2 (12): a011627. DOI:10.1101/cshperspect.a011627. ISSN 2157-1422. PMC 3543078. PMID 23209182.
К группе
гемопротеинов относятся гемоглобин и его производные, миогло-бин,
хлорофиллсодержащие белки и ферменты (вся цитохромная система, каталаза и
пероксидаза). Все они содержат в качестве небелкового компонента структурно
сходные железо- (или магний)порфирины, но различные по составу и структуре
белки, обеспечивая тем самым разнообразие их биологических функций. Далее более
подробно рассмотрено химическое строение гемоглобина, наиболее важного для
жизнедеятельности человека и животных соединения.
Гемоглобин в
качестве белкового компонента содержит глобин, а небелкового – гем. Видовые
различия гемоглобина обусловлены глобином, в то время как гем одинаков у всех
видов гемоглобина.
Основу
структуры простетической группы большинства гемосодержащих белков составляет
порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного
соединения – порфирина. Последний состоит из четырех замещенных пирролов:
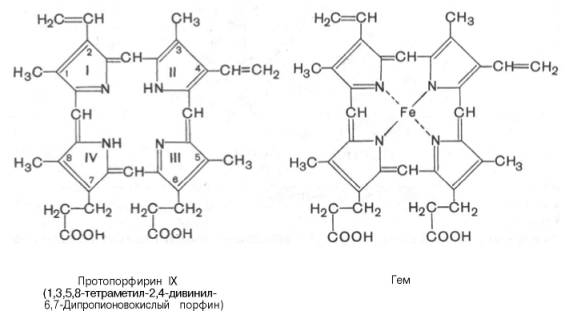
соединенных
между собой метиновыми мостиками (—СН=). Незамещенный порфирин называется
порфином. В молекуле гема порфин представлен в виде протопорфирина IX,
содержащего четыре метильные группы (—СН3), две винильные группы
(—СН=СН2) и два остатка пропионовой кислоты. Протопорфирин,
присоединяя железо, превращается в гем.
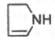
Из формулы
видно, что железо связано с двумя атомами азота молекулы протопорфирина
ковалентно и с двумя другими – координационными связями, обозначенными
пунктирными линиями. В зависимости от химической природы групп, находящихся в
боковой цепи, порфирины классифицируют на этио-, мезо-, копро- и
протопорфирины. Последние наиболее распространены в природе. Из возможных 15
изомеров протопорфиринов благодаря наличию трех разных заместителей самым
распространенным оказался протопорфирин IX.
Гем в виде
гем-порфирина является простетической группой не только гемоглобина и его
производных, но и миоглобина, каталазы, пероксидазы и цитохромов b, с и c1
(см. главу 9); в то же время в цитохромах а и
a3, входящих
в состав интегрального комплекса, названного цитохромокси-дазой, содержится гем
а, называемый также формилпорфирином:
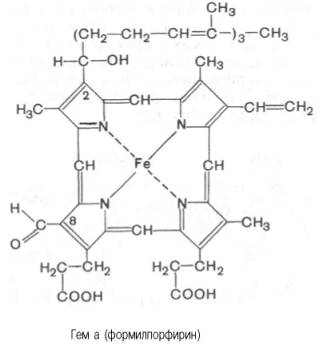
Гем а вместо
метильной группы содержит формильный остаток (в 8-м положении) и вместо одной
винильной группы (во 2-м положении) – изопре-ноидную цепь. Железо своими
четырьмя связями образует комплекс с порфирином, а оставшиеся 5-я и 6-я
координационные связи железа в молекулах гемоглобина и цитохромов связываются с
белковыми компонентами по-разному. В частности, в гемоглобинах (и миоглобине)
благодаря 5-й координационной связи железо соединяется с атомом азота
имидазольной группы гистидина белковой молекулы. Шестая координационная связь
железа предназначена для присоединения кислорода (с образованием
оксигемоглобина и оксимиоглобина) или других лигандов: СО, цианидов и др. (рис.
2.1). В цитохромах, напротив, и 5-я, и 6-я координационные связи железа
соединены с остатками гистидина и метио-нина (в цитохроме с обе винильные группы соединены еще и с
остатками цистеина) белковой молекулы. Этим, вероятнее всего, могут быть объяснены
функции железа в гемоглобине, валентность которого не изменяется при
присоединении кислорода (в отличие от валентности железа в цитохромах): в
гемоглобине железо остается двухвалентным независимо от присоединения или
отдачи кислорода.
Структурная
организация гемоглобина (и миоглобина) была описана в главе 1. Дж. Кендрью и М.
Перутц расшифровали конформацию этих молекул (Нобелевская премия 1962 г.).
Дыхательная функция гемоглобина крови подробно рассматривается в курсе
физиологии. Здесь следует указать на уникальную роль гемоглобина в траспорте
кислорода от легких к тканям и диоксида углерода от тканей к легким. Это
элементарное проявление жизни – дыхание, хотя и выглядит простым, основано на
взаимодействии многих типов атомов в гигантской молекуле гемоглобина.
Подсчитано, что в одном эритроците содержится около 340000000 молекул
гемоглобина, каждая из которых состоит примерно из 103атомов С, Н,
О, N, S и 4 атомов железа.
Атом железа
расположен в центре гема-пигмента, придающего крови характерный красный цвет.
Каждая из 4 молекул гема «обернута» одной полипептидной цепью. В молекуле
гемоглобина взрослого человека HbА (от англ. adult –
взрослый) содержатся четыре полипептидные цепи, которые вместе составляют
белковую часть молекулы – глобин. Две из них, называемые α-цепями, имеют
одинаковую первичную структуру и по 141 аминокислотному остатку. Две другие,
обозначаемые β-цепями, также идентично построены и содержат по 146
аминокислотных остатков. Таким образом, вся молекула белковой части гемоглобина
состоит из 574 аминокислот. Во многих положениях α- и β-цепи
содержат разные аминокислотные последовательности, хотя и имеют почти
одинаковые пространственные структуры. Получены доказательства, что в структуре
гемоглобинов более 20 видов животных 9 аминокислот в последовательности
оказались одинаковыми, консервативными (инвариантными), определяющими функции
гемоглобинов; некоторые из них находятся вблизи гема, в составе участка
связывания с кислородом, другие – в составе неполярной внутренней структуры
глобулы.
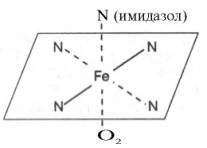
Рис. 2.1. Координационные
связи атома железа в молекуле гема. Все 4 связи с атомами азота пиррольных
колец расположены в одной плоскости, 5-я и 6-я координационные связи (с атомом
азота имидазольного кольца гистидина и с кислородом соответственно) – по разные
стороны перпендикулярно к этой плоскости.
В дополнение
к основному гемоглобину HbA1в крови
взрослого человека доказано существование мигрирующего с меньшей скоростью при
электрофорезе гемоглобина НbА2, также состоящего из 4 субъединиц:
двух α-цепей и двух δ-цепей. На долю НbА2 приходится около
2,5% от всего гемоглобина. Известен, кроме того, фетальный гемоглобин
(гемоглобин новорожденных), обозначаемый HbF и состоящий
из двух α-цепей и двух γ-цепей. Фетальный гемоглобин отличается от HbA1не только составом аминокислот, но и физико-химическими свойствами:
спектральным показателем, электрофоретической подвижностью, устойчивостью к
щелочной денатурации и др. Кровь новорожденного содержит до 80% HbF, но
к концу 1-го года жизни он почти целиком заменяется на НbА (все же в крови
взрослого человека открывается до 1,5% HbF от общего
количества гемоглобина). Последовательность аминокислот в γ- и
δ-цепях гемогло-бинов окончательно не расшифрована.
Установление
первичной структуры субъединиц молекулы гемоглобина стимулировало исследования
по расшифровке структуры так называемых аномальных гемоглобинов. В крови
человека в общей сложности открыто около 150 различных типов мутантных
гемоглобинов. Появляются мутантные формы гемоглобинов в крови вследствие мутации
генов. Обычно мутации делят на 3 класса в соответствии с топографией
измененного участка молекулы. Если замена аминокислоты происходит на
поверхности молекулы гемоглобина, то это мутация первого класса; подобные
мутации обычно не сопровождаются развитием тяжелой патологии, и болезнь
протекает бессимптомно; исключение составляет серповидно-клеточная анемия. При
замене аминокислоты вблизи гема нарушается связывание кислорода –
это мутация второго класса, сопровождающаяся развитием болезни. И наконец, если
замена происходит во внутреннем участке молекулы гемоглобина, говорят о третьем
классе мутации; подобные мутации приводят к нарушению пространственной
структуры и соответственно функции гемоглобина.
Аномальные
гемоглобины, различающиеся по форме, химическому составу и величине заряда,
были выделены при помощи электрофореза и хроматографии. Передающиеся по
наследству изменения чаще всего являются результатом мутации единственного
триплета, приводящей к замене одной какой-либо аминокислоты в полипептидных
цепях молекулы гемоглобина на другую. В большинстве случаев происходит замена
кислой аминокислоты на основную или нейтральную (табл. 2.1). Поскольку это
замещение осуществляется в обеих полипептидных цепях одной из пар (α или
β), образовавшийся аномальный гемоглобин будет отличаться от
нормального величиной заряда и соответственно электрофоретической подвижностью.

В табл. 2.1
представлены некоторые типы аномальных гемоглобинов, составы их полипептидных
цепей с указанием известной или вероятной локализации замены либо в α-, либо
в β-цепях. Замены необычной аминокислотой в аномальных гемоглобинах имеют
место как в α-, так и в β-цепях. Исключение
составляет гемоглобин Н, все 4 полипептида которого представлены β-цепями,
идентичными по структуре β-цепям нормального гемоглобина A1.
Следует
указать, что некоторые мутации, вызывающие существенное изменение структуры и
соответственно функции гемоглобина, оказываются летальными, и индивидуумы с
подобным гемоглобином умирают в раннем возрасте. Однако при ряде мутаций замена
аминокислот не вызывает заметного изменения функции гемоглобина, в этих случаях
болезнь протекает бессимптомно.
Болезни
гемоглобинов (их насчитывают более 200) называют гемогло-бинозами. Принято делить их на гемоглобинопатии, в
основе развития которых лежит
наследственное изменение
структуры какой-либо цепи нормального
гемоглобина (часто их относят также к «молекулярным болезням»), и талассемии,
обусловленные наследственным нарушением синтеза какой-либо нормальной цепи
гемоглобина. Различают также же-лезодефицитные анемии.
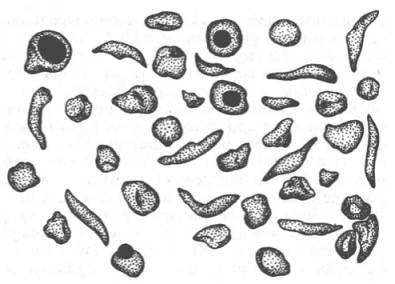
Рис. 2.2. Нормальные
и серповидные эритроциты.
Классическим
примером наследственной гемоглобинопатии является серповидно-клеточная анемия,
широко распространенная в странах Южной Америки, Африки и Юго-Восточной Азии.
При этой патологии эритроциты в условиях низкого парциального давления
кислорода принимают форму серпа (рис. 2.2). Гемоглобин S, как
показали Л. Полинг и др., отличается рядом свойств от нормального гемоглобина:
в частности, после отдачи кислорода в тканях он превращается в плохо
растворимую дез-окси-форму и начинает выпадать в осадок в виде веретенообразных
кристаллоидов, названных тактоидами. Последние деформируют клетку и приводят к
массивному гемолизу. Болезнь протекает остро, и дети, гомозиготные по
мутантному гену, часто умирают в раннем возрасте.
Химический
дефект при серповидно-клеточной анемии был раскрыт В. Ингремом и сводится к
замене единственной аминокислоты, а именно глутаминовой, в 6-м положении с
N-конца на валин в β-цепях молекулы гемоглобина HbS (см.
табл. 2.1, рис. 2.2). Это результат мутации в молекуле ДНК, кодирующей синтез
β-цепи гемоглобина. Все остальные аминокислоты располагаются в той же
последовательности и в таком же количестве, как и в нормальном гемоглобине НbА:
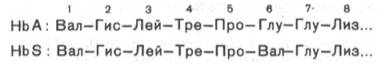
Одной этой
замены оказалось достаточно не только для нарушения формы эритроцита, но и для
развития тяжелой наследственной болезни – серповидно-клеточной анемии.
Талассемии,
строго говоря, не являются гемоглобинопатиями. Это генетически обусловленное
нарушение синтеза одной из нормальных цепей гемоглобина. Если угнетается синтез β-цепей, то
развивается β-талассемия; при
генетическом дефекте синтеза α-цепей развивается α-талас-семия. При
β-талассемии в крови наряду с HbA1появляется
до 15% НbА2 и резко повышается содержание HbF – до 15–60%.
Болезнь характеризуется гиперплазией и разрушением костного мозга, поражением
печени, селезенки, деформацией черепа и сопровождается тяжелой гемолитической
анемией. Эритроциты при талассемии приобретают мишеневидную форму. Механизм
изменения формы эритроцитов объяснить пока не удалось.
В медицинской
практике часто проводят анализ кровяных пигментов, который основан на
исследовании спектроскопических свойств гема гемоглобина, точнее продуктов его
окисления (хлорида гемина и гематина, образующихся соответственно при обработке
гемоглобина уксусной кислотой в присутствии хлорида натрия или разведенными
растворами щелочей). При восстановлении гематина сульфитом аммония в
присутствии глобина образуется производное гемоглобина – гемохромоген,
в котором денатурированный глобин соединен с гемом. Полученный комплекс
имеет характерный спектр поглощения. Этот метод широко применяется в
судебно-медицинской практике при исследовании кровяных пятен.
Из
многообразия производных гемоглобина, представляющих несомненный интерес для
врача, следует прежде всего указать на оксигемоглобин НbО2 –
соединение молекулярного кислорода с гемоглобином. Кислород присоединяется к
каждому гему молекулы гемоглобина при помощи координационных связей железа,
причем присоединение одной молекулы кислорода к тетрамеру облегчает
присоединение второй молекулы, затем третьей и т.д. Поэтому кривая насыщения
гемоглобина кислородом имеет сиг-моидную форму, свидетельствующую о
кооперативности связывания кислорода. Эта кооперативность обеспечивает не
только связывание максимального количества кислорода в легких, но и
освобождение кислорода в периферических тканях; этому способствует также
наличие Н+ и СО2 в тканях с интенсивным обменом. В свою
очередь кислород ускоряет высвобождение СО2 и Н+ в
легочной ткани. Эта аллостерическая зависимость между присоединением Н+,
О2 и СО2 получила название эффекта Бора.
Помимо
кислорода, гемоглобин легко соединяется с другими газами, в частности с СО, NO и
др. Так, при отравлении оксидом углерода гемоглобин прочно с ним связывается с
образованием карбоксигемо-глобина (НbСО). При этом вследствие высокого сродства
к СО гемоглобин теряет способность связывать кислород и наступает смерть от
удушья, недостаточного снабжения тканей кислородом. Однако при быстром
повышении парциального давления кислорода во вдыхаемом воздухе можно добиться
частичного вытеснения СО из связи с гемоглобином и предотвратить летальный
исход.
При
отравлении оксидами азота, парами нитробензола и другими соединениями часть
гемоглобина окисляется в метгемоглобин (НbОН), содержащий трехвалентное железо.
Метгемоглобин также теряет способность к переносу кислорода от легких к тканям,
поэтому при метгемо-глобинемии (вследствие отравления окислителями) в
зависимости от степени отравления может наступить смерть от недостатка
кислорода. Если вовремя оказать помощь, т.е. повысить парциальное давление
кислорода (вдыхание чистого кислорода), то и в этом случае можно вывести
больного из опасного состояния.
Следует
отметить, что самым надежным методом качественного определения различных
производных гемоглобина является исследование их спектров поглощения.
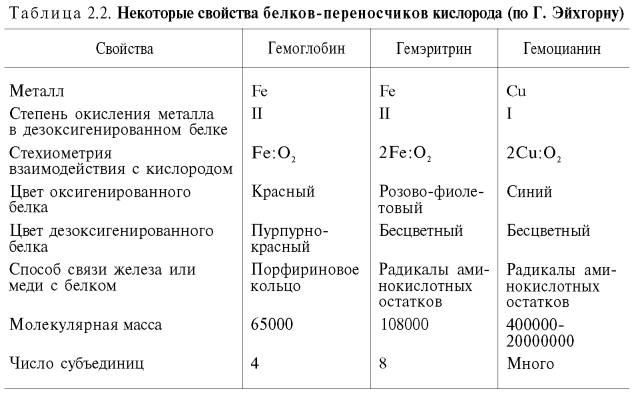
У
беспозвоночных роль переносчика кислорода часто выполняют пигменты негеминовой
природы – гемэритрин и гемоцианин. Они не относятся к гемсодержащим хромопротеинам,
хотя в их названиях содержится корень «гем». Эти белки, как и гемоглобин,
несмотря на то что выполняют одну и ту же функцию, сильно различаются между
собой по молекулярной массе и четвертичной структуре, химической природе
активного центра, характеру связывания железа (гемэритрин) и меди (гемоцианин)
с кислородом и др. (табл. 2.2).
Трансферрины
(сидерофилины) – группа сложных белков, полученных из разных источников и
характеризующихся способностью специфично, прочно и обратимо связывать ионы
железа Fe (III) и других переходных металлов. Наиболее
подробно из этой группы белков изучен трансферрин сыворотки крови. Функция
трансферрина заключается в транспорте ионов железа в ретикулоциты, в которых
осуществляется биосинтез гемоглобина. Система трансферрин–ретикулоцит считается
весьма перспективной для изучения взаимодействия металла с белком и белковой
молекулы с клеткой.
