Грамм гемоглобина мл кислорода

Гемоглобин. Роль гемоглобина в транспорте кислородаОбычно из легких в ткани переносятся эритроцитами в химической связи с гемоглобином около 97% кислорода. Оставшиеся 3% кислорода транспортируются в физической растворенной форме плазмой крови. Таким образом, в нормальных условиях почти весь кислород переносится в ткани, будучи связанным с гемоглобином. а) Обратимая связь кислорода с гемоглобином. Химический состав гемоглобина представлен в отдельных статьях на сайте, где говорилось, что молекула кислорода легко и обратимо связывается с гемом гемоглобина. При высоком PO2, как это бывает в легочных капиллярах, кислород связывается с гемоглобином, а при низком PO2, как в капиллярах тканей, кислород освобождается от связи с гемоглобином. Такой механизм обеспечивает почти весь транспорт кислорода из легких в ткани.
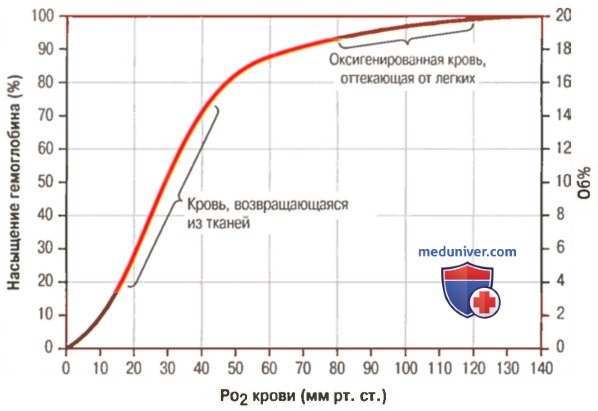
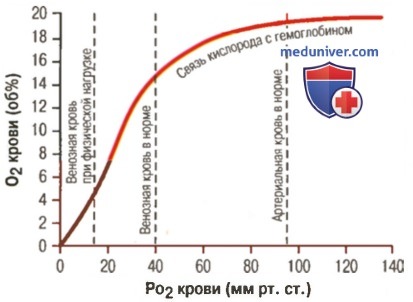
1. Кривая диссоциации оксигемоглобина. На рисунке выше приведена кривая диссоциации оксигемоглобина, демонстрирующая прогрессивный прирост процентной доли оксигемоглобина (процента насыщения гемоглобина кислородом) при увеличении PO2 в крови. В крови, покидающей легкие и входящей в системные артерии, напряжение О2 обычно составляет примерно 95 мм рт. ст., и на кривой диссоциации видно, что насыщение системной артериальной крови кислородом составляет 97%. В нормальной возвращающейся из периферических тканей венозной крови напряжение О2 составляет около 40 мм рт. ст. и 75% — насыщение гемоглобина кислородом. 2. Максимальное количество кислорода, которое может находиться в связи с гемоглобином крови. В 100 мл крови здорового человека содержится около 15 г гемоглобина, и каждый грамм гемоглобина может связать максимально 1,34 мл кислорода (химически чистый гемоглобин может связать 1,39 мл кислорода, но примеси типа метгемоглобина снижают это количество). Итак, 15×1,34 = 20,1, значит, в среднем содержащееся в 100 мл количество гемоглобина при 100% насыщении может связать около 20 мл кислорода. Обычно это обозначают как 20 об% (объемных процентов). Кривая диссоциации оксигемоглобина может строиться не от процентного насыщения гемоглобина кислородом, а от количества содержания объемных процентов кислорода. 3. Количество кислорода, высвобождаемого гемоглобином во время прохождения артериальной крови через ткани. В обычных условиях в системной артериальной крови, насыщенной кислородом на 97%, общее количество связанного с гемоглобином кислорода составляет около 19,4 мл на 100 мл крови (для облегчения понимания просим вас изучить рисунок ниже). При прохождении через капилляры ткани этот объем снижается до 14,4 мл (PO2 — 40 мм рт. ст., насыщение гемоглобина — 75%). Итак, в нормальных условиях каждые 100 мл крови доставляют от легких к тканям около 5 мл кислорода.
4. Транспорт кислорода при тяжелой физической нагрузке. При тяжелой физической работе мышечные клетки потребляют кислород с повышенной скоростью, что может привести к падению PO2 в интерстициальной жидкости мышцы от нормального уровня 40 мм рт. ст. до 15 мм рт. ст. При таком низком парциальном давлении в каждых 100 мл крови остается только 4,4 мл связанного с гемоглобином кислорода. В этом случае каждые 100 мл протекающей крови отдают тканям 19,4 – 4,4, или 15 мл кислорода, т.е. каждый объем крови отдает тканям в 3 раза больше кислорода, чем в покое. Вспомните, что у хорошо тренированных бегунов-марафонцев сердечный выброс может увеличиться в 6-7 раз, чем при покое. Если умножить это увеличение сердечного выброса (6-7 раз) на увеличение высвобождения кислорода в тканях каждым объемом крови (3 раза), получается, что к тканям было доставлено в 20 раз больше кислорода, чем в покое. Далее в этой главе Вы узнаете о существовании других факторов, улучшающих доставку кислорода в ткани во время физической нагрузки, поэтому даже при очень напряженной физической работе наблюдается только очень небольшое снижение PO2 в мышечной ткани. – Также рекомендуем “Коэффициент использования кислорода. Сохранение постоянства кислорода в тканях” Оглавление темы “Кислород и его доставка в организме”: |
Источник
Оглавление темы “Вентиляция легких. Перфузия легких кровью.”:
1. Вентиляция легких. Вентиляция кровью легких. Физиологическое мертвое пространство. Альвеолярная вентиляция.
2. Перфузия легких кровью. Влияние гравитации на вентиляцию легких. Влияние гравитации на перфузию легких кровью.
3. Коэффициент вентиляционно-перфузионных отношений в легких. Газообмен в легких.
4. Состав альвеолярного воздуха. Газовый состав альвеолярного воздуха.
5. Напряжение газов в крови капилляров легких. Скорость диффузии кислорода и углекислого газа в легких. Уравнение Фика.
6. Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
7. Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.
8. Углекислый газ. Транспорт углекислого газа.
9. Роль эритроцитов в транспорте углекислого газа. Эффект Холдена..
10. Регуляция дыхания. Регуляция вентиляции легких.
Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл • л-1 • кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
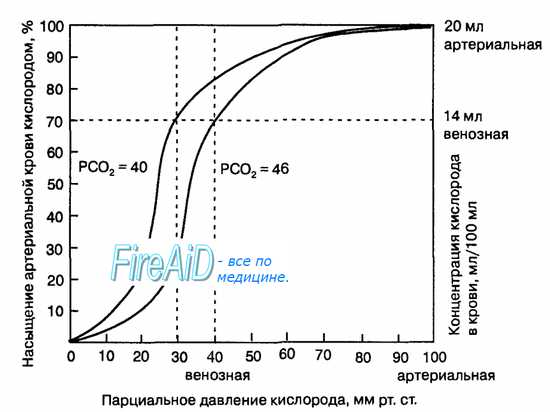
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
– Также рекомендуем “Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.”
Источник
Дыхательная или респираторная система. Гемоглобин и углекислый газ
Мы, рискуя навлечь справедливые нарекания, позволим себе заявить, что основными системами, обеспечивающими поддержания жизнедеятельности больного при критических состояниях, являются системы органов дыхания и кровообращения. Это утверждение в аспекте тематики настоящей монографии становится обоснованным, если учесть, что при современном уровне интенсивной терапии критических состояний основное внимание реаниматолога направлено на поддержание или протезирование функций именно данных систем.
Поэтому-то большинство методов мониторинга при критических состояниях направлены на регистрацию функциональных параметров газообмена и гемодинамики.
Далее мы намеренно ограничили изложение физиологии и патологии дыхания и кровообращения только освещением вопросов, связанных с газообменной функцией организма, поскольку именно она является основным объектом мониторинга критических состояний. Поэтому основное внимание будет посвящено проблеме газотранспортной функции органов дыхания, крови и насосной функции сердца, обеспечивающих транспорт газов. В связи с этим изложение материала было разделено на две части.
Первая глава посвящена физиологии и патологии газотранспортной функции крови и системы органов дыхания, вторая — насосной функции сердца — основному механизму, обеспечивающему транспорт газов к органам и тканям.
Детальное описание физиологии и патологии дыхания и кровообращения можно найти в известных монографиях отечественных и зарубежных авторов, посвященных тщательному анализу основных вопросов этой проблемы и на нашем сайте МедУнивер.

В плазме крови при нормальном атмосферном давлении кислород растворен в минимальных количествах, всего в 0,03% (9 мл на 3 литра циркулирующей крови).
В эритроцитах крови находится сложное белковое вещество — гемоглобин, которое, в свое время, великий английский физиолог Дж.Баркрофт назвал «чудесным». Своеобразие этого вещества состоит в том, что, во-первых, оно обладает повышенным сродством к кислороду (1 грамм гемоглобина способен присоединить 1,34-1,37 мл кислорода) и при нормальном содержании гемоглобина 150 г 1 л крови может связать около 200 мл кислорода.
Во-вторых, степень сродства гемоглобина к кислороду меняется в зависимости от парциального давления (напряжения) кислорода. Чем меньше напряжение кислорода, тем больше к нему сродство гемоглобина и тем быстрее он присоединяет кислород.
Эта особенность гемоглобина имеет большой физиологический смысл. При напряжении кислорода в 27 мм Нд гемоглобин насыщается кислородом на 50%, а при напряжении кислорода 60-70 мм Нд гемоглобин почти полностью насыщен кислородом. Отсюда следует, что транспорт кислорода полностью обеспечивается даже в случае 30-40% дефицита поступающего в организм кислорода. Поистине Дж. Баркрофт был прав, назвав гемоглобин чудесным веществом.
Углекислый газ находится в плазме крови в виде раствора и в химической связи в виде легко диссоциирующего гидрокарбоната натрия. Растворимость углекислоты приблизительно в 20 раз больше, чем у кислорода, в связи с этим ее транспорт осуществляется в основном путем диффузии из плазмы.
Однако при некоторых состояниях, сопровождающихся недонасыщением крови кислородом (гипоксемия), традиционный транспорт углекислого газа может дополняться и транспортом в связанном состоянии с гемоглобином.
– Также рекомендуем “Транспорт газов в организме. Особенности дыхательной системы человека”
Оглавление темы “Контроль обмена газов у пациента”:
1. Метаболизм в организме. Значение кислорода
2. Дыхательная или респираторная система. Гемоглобин и углекислый газ
3. Транспорт газов в организме. Особенности дыхательной системы человека
4. Механизмы транспорта газов. Физиология дыхания – кислородный каскад
5. Диффузия газов. Транспорт кислорода из атмосферы в альвеолы
6. Влияние ЦНС на обмен кислорода. Градиент напряжения кислорода
7. Нарушение напряжения кислорода в альвеолярном газе. Факторы влияющие на напряжение кислорода в альвеолах
8. Транспорт кислорода из альвеол в капилляры. Функции аэрона
9. Диспропорции транспорта кислорода в легких. Нарушения функций аэрона
10. Нарушение альвеоло-артериального градиента кислорода. Транспорт кислорода к тканям
Источник
Из сопоставления сатурационной кривой для кислорода (рис. 10.8) и кривой диссоциации оксигемоглобина (рис. 10.10) видно, что при 100% насыщении гемоглобина кислородом объемная концентрация кислорода в крови составляет (в норме!) около 20 об%. Рассмотрим, каким образом получается такое значение.
Поскольку подавляющая часть кислорода в крови приходится на химически связанный с гемоглобином кислород, объемная концентрация кислорода в крови зависит от:
¾ концентрациигемоглобина;
¾ кислородной емкости гемоглобина, то есть максимального количества кислорода, которое может связать 1 г гемоглобина при полном насыщении;
¾ степени насыщениягемоглобина кислородом:
cO2 = Hb% ´ КЕHb ´ sO2, (16)
где cO2 — объемная концентрация кислорода; Hb% — концентрация гемоглобина в крови; КЕHb — кислородная емкость гемоглобина; sO2 — насыщениегемоглобина кислородом.
В идеальном случае для артериальной крови:
¾ Hb% равна 15 г/100 мл;
¾ КЕHb равна 1,34 — один грамм гемоглобина при полном насыщении способен связать до 1,34 мл кислорода;
¾ sO2 равно около 100%.
Итак:
cO2 = 15 (г Hb/100 мл крови) ´ 1,34 (мл O2/г Hb) ´ 100%= 20 (мл O2/100 мл крови).(17)
Парциальные давления и объемные концентрации кислорода в артериальной и венозной крови приведены в табл. 10.1.
Углекислый газ
Этот газ переносится в нескольких формах, или транспортных фракциях:
¾ физически растворенный CO2;
¾ CO2, соединенный с гемоглобином (карбогемоглобин);
¾ бикарбонат (HCO3–).
Для того чтобы мог образоваться бикарбонат, CO2 должен сначала прореагировать с H2O с образованием угольной кислоты. Эта реакция катализируется ферментом карбоангидразой,находящейся в эритроцитах. Таким образом, для образования бикарбоната CO2 должен сначала войти в эритроцит. Поскольку гемоглобин имеется только в эритроцитах, для образования карбогемоглобинаCO2 также должен войти в эритроцит. Образующийся в эритроцитах бикарбонат должен выйти в плазму, а для сохранения электронейтральности взамен в эритроцит должен поступить другой анион — основной анион плазмы Cl–. Это осуществляет мембранный белок — хлор-бикарбонатный обменник.
Итак, образование основных транспортных фракций CO2 происходит в эритроците, и для этого эритроцит обладает следующими свойствами:
¾ в цитоплазме эритроцита имеется карбоангидраза;
¾ в цитоплазме эритроцита имеется гемоглобин;
¾ мембрана эритроцита (как и любые другие мембраны) свободно пропускает CO2;
¾ в мембране эритроцита имеется хлор-бикарбонатный обменник.
Образование транспортных фракций CO2
Последовательность событий при образовании этих фракций следующая (рис. 10.11).
1. Выделяемый тканями CO2 проходит через стенку капилляра по градиенту его парциального давления; образуется первая фракция — физически растворенный CO2.
2. Из плазмы CO2 проходит через мембрану эритроцита.
3. В эритроците CO2 соединяется с гемоглобином; образуется вторая фракция — карбогемоглобин.
4. В эритроците CO2 под действием карбоангидразы соединяется с водой; образуется угольная кислота.
5. Угольная кислота распадается до H+ и HCO3–; образуется третья фракция — бикарбонат.
6. Бикарбонат выходит из эритроцита в обмен на Cl– (с помощью хлор-бикарбонатного обменника).
7. Образующиеся на этапе 5 протоны (H+) связываются с гемоглобином, так как он обладает буферными свойствами (гл. 9).
Сатурационная кривая для CO2
Основная транспортная фракция CO2 — бикарбонат, который может образовываться почти бесконечно (до тех пор, пока CO2 не свяжется со всеми молекулами воды, что в физиологических условиях невозможно). Отсюда:
¾ объемная концентрация CO2 гораздо выше, чем O2 (табл. 10.1);
¾ сатурационная кривая для CO2 не характеризуется насыщением (не имеет горизонтального участка).
Эта кривая приведена на рис. 10.12.
Фракция карбогемоглобина придает сатурационной кривой для CO2 важное свойство (аналогичное соответствующему свойству для кислорода): при снижении парциального давления кислорода (pO2) кривые сдвигаются вверх и влево, то есть сродство гемоглобина к CO2 увеличивается. Поскольку pO2 снижено в активно работающих тканях, то, проходя через такие ткани, кровь дополнительно связывает интенсивно выделяемый ими CO2.
Парциальные давления и объемные концентрации углекислого газа в артериальной и венозной крови приведены в табл. 10.1.
Источник