Кривая насыщения кислородом гемоглобина

Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 10 июля 2013;
проверки требуют 29 правок.
Эффект Вериго — Бора (синонимы — эффект Вериго, эффект Бора) — зависимость степени диссоциации оксигемоглобина от величины парциального давления углекислоты в альвеолярном воздухе и крови, при снижении которого сродство кислорода к гемоглобину повышается, что затрудняет переход кислорода из капилляров в ткани. Эффект этот был открыт независимо друг от друга Б. Ф. Вериго в 1892 году[1] и датским физиологом К. Бором в 1904 году[2].
История открытия[править | править код]
Основу разработки проблемы гипоксии заложил русский учёный-физиолог И. М. Сеченов фундаментальными работами по физиологии дыхания и газообменной функции крови. Большое значение имеют также исследования русского физиолога Б. Ф. Вериго по физиологии газообмена в лёгких и тканях. Опираясь на идеи Сеченова о сложных формах взаимодействия между диоксидом углерода и кислородом в крови (Вериго работал в лабораториях Сеченова, И. Р. Тарханова и И. И. Мечникова), он впервые установил зависимость степени диссоциации оксигемоглобина от величины парциального давления углекислоты в крови.
Биохимический механизм[править | править код]
Эффект Вериго — Бора: влияние pH на кривую насыщения гемоглобина кислородом
В клетках периферических тканей органическое топливо окисляется в митохондриях с использованием кислорода (клеточное дыхание), доставляемого гемоглобином из лёгких; при этом в качестве продуктов образуются углекислый газ, вода и другие соединения. Образование углекислого газа в тканях одновременно приводит и к повышению концентрации ионов H+ (то есть к понижению pH), поскольку при гидратации CO2 образуется H2CO3 — слабая угольная кислота, диссоциирующая на ионы H+ и бикарбонат-ионы:
H2CO3 ⇄ H+ + HCO3-.
Гемоглобин переносит значительную долю (около 20 %) общего количества CO2 и ионов H+, образующихся в тканях и поступающих в лёгкие и почки, обеспечивающих выделение этих продуктов.
За много лет до открытия этого механизма было обнаружено, что на связывание кислорода гемоглобином очень сильное влияние оказывает pH и концентрация CO2: при присоединении CO2 и ионов H+ способность гемоглобина связывать O2 снижается. Действительно, в периферических тканях с относительно низким значением pH и высокой концентрацией CO2 сродство гемоглобина к кислороду падает. И наоборот, в лёгочных капиллярах выделение CO2 и сопутствующее ему повышение pH крови приводит к увеличению сродства гемоглобина к кислороду. Это влияние величины pH и концентрации CO2 на связывание и освобождение O2 гемоглобином и называют эффектом Вериго — Бора.
Реакция связывания кислорода гемоглобином в виде
Hb + O2 ⇄ HbO2
в действительности отражает неполную картину, поскольку не учитывает дополнительные лиганды H+ и CO2.
Чтобы объяснить влияние концентрации ионов H+ на связывание кислорода, следует записать эту реакцию в иной форме:
HHb+ + O2 ⇄ HbO2 + H+,
где HHb+ — протонированная форма гемоглобина. Из этого уравнения следует, что кривая насыщения гемоглобина кислородом зависит от концентрации ионов H+. Гемоглобин связывает и O2, и ионы H+, но между этими двумя процессами существует обратная зависимость. Если парциальное давление кислорода велико (что наблюдается, например, в лёгких), то гемоглобин связывает его, освобождая при этом ионы H+. При низком парциальном давлении кислорода (что имеет место в тканях) связываться с гемоглобином будут ионы H+.
Эффективность связывания гемоглобином углекислого газа (с образованием карбаминогемоглобина, чаще именуемого карбгемоглобином[3]) находится в обратной зависимости от связывания кислорода. В тканях часть избыточного CO2 связывается с гемоглобином, сродство последнего к O2 снижается, и происходит высвобождение кислорода. В лёгких же связывается избыток O2 воздуха, тем самым сродство гемоглобина к CO2 уменьшается, и CO2 выделяется в альвеолярный воздух, способствуя лёгкому закислению крови за счёт ионов H+, возникающих при диссоциации угольной кислоты (см. выше).
Описанные зависимости делают молекулу гемоглобина великолепно приспособленной к осуществлению совместного переноса эритроцитами кислорода, углекислого газа и ионов H+[4].
См. также[править | править код]
- Эффект Холдейна (открыт Джоном Скоттом Холдейном)
Примечания[править | править код]
- ↑ Verigo B. F. Zur Frage über die Wirkung des Sauerstoff auf die Kohlensäureausscheidung in den Lungen // Archiv für die gesammte Physiologie des Menschen und der Thiere : статья. — 1892. — № 51. — С. 321—361.
- ↑ Chr. Bohr, K. Hasselbalch, and August Krogh. Concerning a Biologically Important Relationship — The Influence of the Carbon Dioxide Content of Blood on its Oxygen Binding // Skand. Arch. Physiol. : статья. — 1904. — № 16. — С. 401—412.
- ↑ Карбгемоглобин // Биологический энциклопедический словарь (рус.).
- ↑ Ленинджер А. Основы биохимии: В 3-х томах. Том 1. — М.: Мир, 1985. — С. 208—210. — 367 с.
Литература[править | править код]
- Вериго эффект // Большой медицинский словарь (рус.). — 2000.
Источник
Несмотря на то, что исследование кислотно-основного состояния, строго говоря, подразумевает исследование только величины pH (концентрации ионов H+), в реальности в него также включается исследование физиологически важных газов, присутствующих в крови – O2 и CO2. Анализ газов показывает эффективность газообмена по величинам парциальных давлений – pO2 и pCO2.
Через альвеолярную мембрану молекулы любых газов перемещаются диффузно по градиенту концентрации. Молекулы O2 атмосферного воздуха поступают из альвеол в кровь, а молекулы CO2 из крови в альвеолы до тех пор пока их парциальные давления не выровняются.
Величина парциального давления – это процентная доля газа в общем объеме.
Углекислый газ
Концентрация СО2 в альволярном воздухе столь низка, а в крови столь высока, что диффузия этого газа в альвеолы чрезвычайно эффективна и скорость его удаления зависит только от альвеолярной вентиляции – общего объема воздуха, транспортируемого в минуту между альвеолами и атмосферой (“скорости выдувания”).
Следовательно,
- при усиленной вентиляции легких углекислый газ быстро выводится, и показатель pCO2в крови снижается. Это означает потерю организмом угольной кислоты (ионов H+), что является причиной защелачивания крови – алкалоза, называемого дыхательным или респираторным.
- при недостаточной альвеолярной вентиляции величина рСО2 повышается, что свидетельствует о недостаточном его удалении и накоплении H2CO3. Иными словами, повышение в крови показателя рСО2 является причиной дыхательного ацидоза.
Увеличенное pCO2 (гиперкапния) всегда свидетельствует о снижении альвеолярной вентиляции.
Кислород
Вопросы, связанные с оксигенацией крови и транспортом кислорода более сложны. Связано это с тем, что в виде свободных (растворенных) молекул O2 находится лишь небольшая доля общего кислорода крови. Основная часть кислорода связана с гемоглобином (оксигемоглобин) и истинное содержание кислорода зависит от двух дополнительных параметров – концентрации Hb и насыщения (сатурации) гемоглобина кислородом.
Оксигемоглобин
Оксигемоглобин (HbО2) – процентное содержание в крови, является отношением фракции оксигемоглобина (HbО2) к сумме всех фракций (общему гемоглобину).
Насыщение гемоглобина кислородом
Насыщение гемоглобина кислородом (HbOSAT, SО2), представляет собой отношение фракции оксигенированного гемоглобина к тому количеству гемоглобина в крови, который способен транспортировать О2.
Отличия между двумя показателями HbО2 и HbOSAT заключаются в том, что у пациентов возможно наличие в крови такой формы гемоглобина, которая не способна акцептировать О2 (Hb‑CO, metHb, сульфоHb). Но так как большинство больных не имеют в крови повышенного содержания этих форм гемоглобина, значения HbО2 и SО2 обычно очень близки.
Например, если при отравлении нитритами количество metHb составляет 15%, тогда величина HbО2 никогда не сможет превысить 85%, но насыщение (HbOsat) может быть различно – от максимума (HbOsat=95-98%) при полном насыщении до низких величин при отсутствии кислорода.
Показатель насыщения кислородом показывает процент доступных мест связывания на гемоглобине.
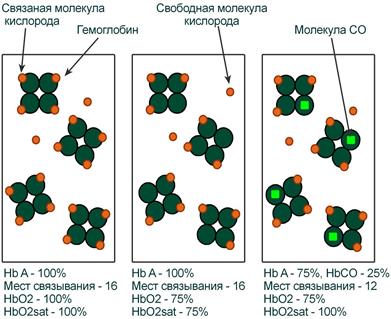
Иллюстрация понятий оксигемоглобина (HbO2) и насыщения гемоглобина (HbO2sat)
Парциальное давление кислорода (pO2)
Парциальное давление O2 выступает как движущая сила, приводящая к насыщению гемоглобина кислородом. И хотя, как правило, чем выше pO2 тем выше HbOsat, эта зависимость не является линейной.
Кривая диссоциации гемоглобина в норме и
при изменении pH и концентрации 2,3-дифосфоглицерата
В центральной части кривой насыщения (или кривой диссоциации) гемоглобина малейшие сдвиги pO2 приводят к резким изменениям насыщения гемоглобина. И наоборот, при высоком pO2 (80-90-100 мм рт.ст) кривая становится плоской, насыщение гемоглобина мало зависит от колебаний кислорода в плазме.
Сдвиг влево происходит при защелачивании и снижении концентрации 2,3-дифосфоглицерата и сигнализирует об увеличении сродства кислорода к гемоглобину (в легких). Сдвиг вправо – это снижение сродства кислорода к гемоглобину (в тканях), обеспечивается закислением среды и накоплением 2,3-дифосфоглицерата.
Показатель pO2 не отражает содержание кислорода в цельной крови! Но хотя pO2 и не показывает общее количество кислорода в крови, но это общее количество зависит от pO2 через показатель сатурации гемоглобина.
В свою очередь имеются факторы, влияющие на величину pO2:
1. Альвеолярная вентиляция. Хотя она влияет как на pO2 так и на pCO2, но доля кислорода в альвеолах при гипервентиляции может лишь слегка увеличиться, приближаясь к pO2 атмосферного воздуха, при гиповентиляции – стремительно падает, вытесняясь поступающим из крови CO2. В то же время доля CO2 в альвеолах быстро снижается при усиленной вентиляции.
2. Вентиляционно-перфузионное соотношение, определяется тем, что
- не вся кровь, притекающая к легким, соприкасается с хорошо вентилируемыми альвеолами (спадение альвеол, уплотнение стенки).
- не все хорошо вентилируемые альвеолы получают достаточно крови (правожелудочковая сердечная недостаточность).
3. Концентрация кислорода во вдыхаемом воздухе (FiO2, fraction of inspired oxygen).
В таблице приведены сравнительные величины концентрации кислорода и углекислого газа в воздухе, крови и тканях.
Необходимо обратить внимание на перепады концентраций кислорода и углекислого газа в крови и альвеолярном воздухе. Важной особенностью является то, что pO2 в альвеолярном воздухе и артериальной крови очень близки, т.е. в обычных условиях глубоким и/или частым дыханием невозможно повысить потребление кислорода и насыщение им гемоглобина. В то же время разность концентраций pCO2 в венозной крови и альвеолярном воздухе позволяет эффективно его удалять при частом дыхании.
| pO2, мм рт.ст. | pCO2, мм рт.ст | |
| Вдыхаемый воздух | 159 | 0,23 |
| Альвеолярный воздух | 105-110 | 40 |
| Артериальная кровь | 83-108 | 35-45 |
| Ткани | 10-20 | 50-60 |
| Венозная кровь | 35-49 | 46-51 |
| Выдыхаемый воздух | 116 | 32 |
Источник
Аллостерическая регуляция насыщения гемоглобина кислородом
Кривая диссоциации кислорода для гемоглобина
Механизм насыщения гемоглобина кислородом
Гемоглобин присоединяет О2 последовательно, по одной молекуле на каждый гем.
В апогемоглобине, благодаря координационной связи с белковой частью, атом железа выступает из плоскости гема в направлении гистидина F8.
Присоединение О2 к шестой координационной связи железа вызывает его перемещение в плоскость гема, за ним перемещаются гистидин F8 и полипептидная цепь, в состав которой он входит.
Происходит изменение конформации текущего протомера и связанных с ним оставшихся протомеров. При этом у протомеров возрастает сродство к кислороду, в результате каждый следующий кислород присоединяется к гемоглобину лучше предыдущего. Четвертая молекула кислорода присоединяется к гемоглобину в 300 раз легче, чем первая молекула. Обратный процесс аналогичен, чем больше О2 отдают протомеры, тем легче идет отщепление последующих молекул О2.
Кооперативность в работе протомеров гемоглобина формирует сигмовидный характер кривой насыщения его кислородом в зависимости от парциального давления кислорода.
S–образная кривая насыщения гемоглобина кислородом имеет важное биологическое значение.
Во-первых, пологий участок S–образной кривой (выше 60 мм.рт.ст.) обеспечивает максимальное насыщение гемоглобина кислородом в легких, даже если концентрация кислорода в альвеолярном воздухе заметно снижена. Например, в альвеолярной крови при РО2=95 мм.рт.ст. гемоглобин насыщается кислородом на 97%, а при РО2=60 мм.рт.ст. – на 90%.
Во-вторых, Крутой наклон среднего участка S–образной кривой (от 10 до 40 мм.рт.ст.) обеспечивает максимальный переход кислорода от гемоглобина к тканям.
В области венозного конца капилляра при РО2 = 40 мм.рт.ст. гемоглобин насыщен кислородом на 73%. При снижении РО2 на 5 мм.рт.ст. насыщение гемоглобина кислородом уменьшается на 7%.
Кроме РО2 на насыщение гемоглобина кислородом влияют и другие факторы, например, рН, температура, давление, концентрация 2,3-ДФГ, РСО2.
Увеличение температуры, присоединение к гемоглобину Н+, 2,3-ДФГ, СО2 уменьшает сродство гемоглобина к кислороду, при этом кривая диссоциации оксигемоглобина сдвигается вправо и гемоглобин легче отдает кислород тканям.
Влияние рН на характер кривой диссоциации оксигемоглобина называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект).
Гемоглобин в дезоксигенерированном состоянии имеет более высокое сродство к протонам, чем оксигемоглобин. Другими словами R – форма (оксигенерированная) является более сильной кислотой, чем Т-форма (дезоксигенерированная). Поэтому когда дезоксигемоглобин в легких присоединяет кислород, происходит переход в R – форму и разрыв некоторых связей, в результате чего и высвобождаются протоны, ответственные за эффект Бора. Наоборот, при высвобождении кислорода образуется Т-структура и разорванные связи между субъединицами должны быть восстановлены, и протоны вновь присоединяются к остаткам гистидина в b – цепях. Таким образом, протонирование гемоглобина снижает его сродство к О2 и увеличивает потребление О2 в ткани.
Эффект Бора имеет важное физиологическое значение. Образующийся в тканях СО2 должен транспортироваться в легкие. Он поступает в эритроциты по градиенту напряжения. В них фермент карбоангидраза превращает его в Н2СО3, который диссоциирует на бикарбонат, ион и протон. Последний сдвигает равновесие влево в уравнении (1).
Hb + 4 O2= Hb (О2)4 + (H+)n
Где n – величина порядка 2; число зависит от целого комплекса параметров, тем самым заставляя Hb О2 отдавать свой кислород.
НСО3- пассивно продвигается через ионный канал по градиенту концентрации в сыворотку.
Продвижение НСО3- не сопровождается перемещением Н+, поскольку нет канала, позволяющего ему пройти через мембрану эритроцитов. Для сохранения ионного равновесия при выходе НСО3- из клетки, Cl- перемещаются внутрь её через тот же ионный канал. Такое двойное перемещение известно как хлоридный сдвиг (сдвиг Хамбургера).
Растворенный НСО3- движется вместе с венозной кровью обратно в легкие. Здесь высвобождение протона из гемоглобина при оксигениции приводит к образованию НСО3- (по принципу Ле-Шателье).
НСО3-+ Н+= Н2СО3-,
что позволяет карбоангидразе образовать СО2.
Разрушение НСО3- в эритроците обуславливает вхождение в него НСО3- из сыворотки, так что в легких происходит обратный хлоридный сдвиг, приводящий к выведению СО2 с выдыхаемым воздухом.
Источник
Этот график – кривая диссоциации оксигемоглобина, или сатурационная кривая, показывает, какая доля гемоглобина в данной крови связана с О2 при том или ином его парциальном давлении, а какая – диссоциирована, т.е. свободна от кислорода. КДО имеет S-образную форму. S-образный вид КДО указывает на кооперативный характер взаимодействия О2 с Hb. При соединении или отдаче одной молекулы О2 отмечаются функциональные изменения остальных субъединиц Hb в тетрамере.Такая её конфигурация, имеет определенный физиологический смысл, заключающийся в том, что оксигенация крови в легких сохраняется на высоком уровне даже при относительно низком альвеолярном pO2, а её деоксигенация существенно изменяется даже при небольшом изменении капиллярно-тканевого градиента pO2. Плато кривой характерно для сатурированной О2 (насыщенной) артериальной крови, а крутая нисходящая часть кривой – венозной, или десатурированной крови в тканях (рис. 2.4.).
В качестве показателя CГK принимается значение pО2, при котором Hb наполовину насыщается О2, обозначаемая p50, равна 26-28 мм рт.ст. При повышении СГК p50 уменьшается, а при снижении – повышается. Традиционно полагают, что сдвиг КДО вправо повышает отдачу кровью кислорода тканям, а влево затрудняет десатурацию крови в микроциркулярном русле.
На кривой выделяют 3 участка: от 0 до 10 мм рт. ст. – прямо пропорциональная зависимость, от 10 до 50 – насыщение очень быстрое, от 60 до 90 – насыщение почти не изменяется. При рО2 в артериальной крови, равном 96 мм рт. ст., % насыщения гемоглобина кислородом составляет 97%, а в венозной крови с рО2, равным 40 мм рт. ст., % насыщения гемоглобина равен 75%.
Значение S-образной формы кривой:
1) Крутой наклон среднего участка, соответствующий напряжениям О2 в тканях (35 мм рт. ст. и ниже), благоприятствует отдаче О2 в них.
2) Пологая часть, КДО соответствующая высоким напряжениям О2, обеспечивает стабильность тканевого рО2в условиях, когда рО2 артериальной крови может уменьшаться: при подъеме в горы или на самолете, при заболеваниях легких, с возрастом. Даже когда альвеолярное рО2 снижается до 60 мм рт. ст., гемоглобин в артериальной крови все еще насыщен О2 на 89%, что только на 8% ниже нормальной 97% сатурации.
3) С другой стороны, когда альвеолярное рО2 увеличивается до 500 мм рт. ст. (при вдыхании воздуха под повышенным давлением, например, на глубине моря или в компрессионных камерах), насыщение гемоглобина кислородом также изменяется мало – максимум на 3%.
Следовательно, при варьировании альвеолярного рО2 от 60 до 500 мм рт. ст. рО2 в тканях изменяется только на несколько мм рт. ст., т.е. гемоглобин буферирует сдвиги тканевого рО2.
В настоящее время принято оценивать положение КДО не по наклону кривой, а по расположению на ней двух точек. Первая соответствует 50%-ному насыщению гемоглобина кислородом – Р50 – напряжение полунасыщения (точка разрядки по Крогу). В норме при рН = 7,4 и t = 37оС, Р50 артериальной крови 26 мм рт. ст. (3,46 кПа). Оно выше у женщин, чем у мужчин. Напряжение разрядки достаточно высоко, что обеспечивает эффективный градиент рО2 между капиллярами и тканями, где рО2 не более 10-15 мм рт.ст. Вторая точка соответствует 95%-ному насыщению гемоглобина кислородом – Р97 – точка зарядки. В норме Р97 артериальной крови 70 мм рт. ст.
КДО может занимать различное положение, характеризующееся смещением ее средней части вправо или влево (рис.2.5. ).
Сдвиги кривой диссоциации
Традиционно полагают, что сдвиг КДО вправо повышает отдачу кровью кислорода тканям, а влево затрудняет десатурацию крови в микроциркулярном русле.
Смещение влево сопровождается снижением Р50 и повышением сродства гемоглобина к О2 (СГК), а вправо – увеличением Р50 и уменьшением СГК.
Сродство кислорода к гемоглобину и способность отдавать О2 в тканях зависит от метаболических потребностей клеток организма и регулируется важнейшими факторами метаболизма тканей, вызывающими смещение кривой диссоциации(рис.2.6.). К этим факторам относятся: концентрация водородных ионов, температура, парциальное напряжение углекислоты и соединение, которое накапливается в эритроцитах – это 2,3-дифосфоглицератфосфат (ДФГ).
Факторы, влияющие на СГК.
По механизму действия их делят на 2 группы: прямые и косвенные.
Прямое влияние оказывают химические вещества, которые могут взаимодействовать с гемоглобином и изменять конформацию его молекулы. Это лиганды (О2, Н+, СО2, органические и неорганические соли).
Косвенное действие оказывают физические факторы (tо, pH), которые изменяют условия взаимодействия Hb с лигандами.
Рассмотрим первую группу факторов:
I. В 1907 C.Bohru, A.Krogh установили зависимость положения КДО от содержания СО2 в крови. Сначала “эффект Бора” связывали только с СО2. Однако затем J.Barcroft (1909) показал, что подобная связь существует и при воздействии других кислот. В настоящее время эффект Бора рассматривается как влияние рН среды на взаимодействие атома Fe2+ с различными лигандами.
При снижении рН (увеличении [Н+]) СГК падает. Поэтому КДО смещается вправо. Уменьшение оксигенации Hb при снижении рН – эффект Рута.
Описанные эффекты объясняются тем, что при изменении рН происходит сдвиг константы диссоциации и конформации групп, связанных с О2 в тетрамере гемоглобина. В результате этого меняется СГК. Различают респираторный и метаболический компоненты эффекта Бора.
Метаболический – связан с изменением рН под влиянием кислых или щелочных продуктов обмена веществ.
Респираторный – связан с влиянием СО2 на КДО. Образующийся при тканевом метаболизме СО2 диффундирует внутрь эритроцитов, где под влиянием карбоангидразы образуется Н2СО3 = Н+ + НСО3־. Образующиеся при ее диссоциации Н+ приводят к снижению рН. Это рН-зависимый эффект СО2. Однако СО2 также образует карбаминовые связи с концевыми группами a и b цепей гемоглобина, способствуя таким образом стабилизации дезоксиформы гемоглобина. Это рН-независимый эффект СО2.
На выраженность эффекта Бора влияют:
1) органические фосфаты – при повышении содержания 2,3-дифосфоглицериновой кислоты (2,3-ДФГ) падает коэффициент Бора, который определяют как D log Р50 / D log рН; 2) температура; 3) насыщение крови О2;
4) электролиты; 5) свойства Hb; 6) пол – коэффициент Бора меньше у мужчин, чем у женщин.
Т.о., эффект Бора – это результат сложного взаимодействия рН, СО2, 2,3-ДФГ и всех других указанных факторов.
Биологический смысл эффекта Бора: он способствует поглощению О2 в легких и отдаче его в тканях.
В легких – поглощение О2 идет одновременно с выделением СО2, поэтому по мере насыщения гемоглобина кислородом КДО сдвигается влево (рис. 2.6), т.е. СГК возрастает. В результате скорость диффузии О2 из альвеол в кровь несколько увеличивается.
В тканях – одновременно с выходом О2 в эритроциты поступает СО2, поэтому КДО смещается вправо, что приводит к снижению СГК и благоприятствует отдаче О2 тканям.
II. В 1967 г. была описана специфическая роль органичнских фосфатов в эритроцитах. 2,3-ДФГ, значительно уступая АТФ в энергетической ценности, играет огромную роль в регуляции транспорта О2. Во-первых, он взаимодействует с b-цепями восстановленного гемоглобина благодаря соответствию зарядов некоторых аминокислотных остатков распределению зарядов в органическом фосфате. После образования связи с b-цепью 2,3-ДФГ закрывает вход в центральный канал молекулы, что уменьшает СГК. Во-вторых, 2,3-ДФГ также изменяет внутриклеточный рН.
На реакцию 2,3-ДФГ и гемоглобина влияют:
1) соотношение Hb/HbО2 – чем оно выше, тем больше связывается 2,3-ДФГ);
2) рН – снижение рН увеличивает сродство гемоглобина к 2,3-ДФГ;
3) рСО2– уменьшение рСО2 повышает сродство гемоглобина к 2,3-ДФГ;
4) tо– возрастание tо препятствует присоединению 2,3-ДФГ к гемоглобину;
5) анионы – могут конкурировать с 2,3-ДФГ за одни и те же центры связывания.
С изменением содержания 2,3-ДФГв эритроцитах связана зависимость СГК от их возраста – молодые эритроциты имеют меньшее СГК, по мере старения оно возрастает, поскольку уровень 2,3-ДФГ уменьшается.
Роль 2,3-ДФГ в качестве регулятора положения КДО заключается не только во влияниии на СГК. Он служит посредником других влияний – рН, рСО2, t, ионного состава на кислородтранспортную функцию крови.
III. Органические и неорганические соли – при увеличении их концентрации КДО сдвигается вправо. Соли влияют через эритроцитарный метаболизм.
К косвенным факторам, как уже отмечалось, относят рН, влияние которого на СГК рассмотрено выше, и tо. Уменьшение tо увеличивает интенсивность образования HbО2, т.е. при прочих равных условиях смещает КДО влево. Повышение tо, напротив, снижает СГК.
Температурный коэффициент, представляющий собой отношение D log рО2 / D tо, зависит от рН, [2,3-ДФГ], насыщения крови кислородом. Он различен у мужчин и женщин. Оксигенация гемоглобина является экзотермической реакцией, соответственно диссоциация оксигемоглобина протекает с поглощением тепла. При повышении температуры крови происходит снижение СГК и его увеличение при снижении температуры. Алкалоз и гипотермия повышают сродство гемоглобина к кислороду. В этих условиях гемоглобин жадно присоединяет к себе кислород в легких и неохотно отдает его тканям.
При ацидозе и гипертермии в эритроцитах сродство гемоглобина к кислороду снижается. При этом насыщение гемоглобина кислородом в легких ухудшается, но отделение кислорода от оксигемоглобина в капиллярах облегчается.
Особый интерес для изучения кислородсвязывающих свойств крови представляет выделение NO клетками эндотелия кровеносных сосудов.
В целом, дыхательный цикл можно рассматривать как механизм транспорта “трех газов”: NO/О2/CО2 . Характерной особенностью NO является способность быстро (менее чем за 5 секунд) диффундировать через мембрану синтезировавшей его клетки в межклеточное пространство и легко (без участия рецепторов) проникать в клетки-мишени. В результате взаимодействия NO и гемоглобина происходит образование его различных NO-форм: метгемоглобин, нитрозилгемоглобин, нитрозогемоглобин, которые играют роль своеобразного аллостерического регулятора функциональной активности гемоглобина на уровне отдельных его тетрамеров. Присутствие этих соединений гемоглобина с NO может по-разному влиять на СГК всей крови: метгемоглобин и нитрозогемоглобин его повышают, а нитрозилгемоглобин снижает. В ходе одного цикла движения эритроцита в сосудистой системе происходят последовательные реакции гемоглобина с NO, модулирующиеся его структурными переходами из R– в Т-состояние. На уровне капилляров малого круга кровообращения это может быть дополнительным механизмом, способствующим оксигенации крови, а на уровне микроциркуляции большого круга – оптимизирующим десатурацию крови, и, соответственно, доставку кислорода в ткани.
Транспорт углекислого газа кровью
Химические реакции связывания СО2 несколько сложнее, чем О2. Это обусловлено тем, что механизмы, отвечающие за транспорт СО2, должны одновременно обеспечивать поддержание кислотно-щелочного равновесия крови и тем самым – внутренней среды организма.
I. Первый способ связывания СО2 – прежде всего происходит гидратация молекул СО2 с образованием Н2СО3
СО2 + Н2О ® Н2СО3
В плазме эта реакция протекает очень медленно. В эритроцитах она ускоряется в 20 тысяч раз, что связано с действием карбоангидразы – фермента, расположенного только внутри клеток. Его существование предполагал еще И.М.Сеченов, но открыт он был в 1932 г. Мелдрумом и Рафтоном.
Следующая реакция в цепи химических превращений СО2 заключается в диссоциации слабой кислоты Н2СО3 на бикарбонат-ионы и Н+.
Н2СО3 ® НСО3־ + Н+
Накопление НСО3־ в эритроцитах приводит к тому, что между его внутренней средой и плазмой крови создается градиент концентрации этого иона. НСО3 могут передвигаться по этому градиенту лишь в том случае, если при этом не будет нарушаться равновесное распределение электрических зарядов. Т.е., одновременно с выходом НСО־3 должен происходить либо выход из эритроцитов одного катиона, либо вход одного аниона(рис.2.7.). Поскольку мембрана эритроцитов практически непроницаема для катионов, но проницаема для небольших анионов, взамен НСО3 в эритроциты поступает Cl-. Этот обменный процесс называется хлоридным сдвигом Хамбургера.
Участие эритроцитов в обмене О2 и СО2 в тканях и в легких
А- между тканями и кровью; Б- между кровью и альвеолярным воздухом
Накопление анионов внутри эритроцитов приводит к повышению осмотического давления, что вызывает поступление в них воды. В результате этого объем эритроцитов в венозной крови больше, чем в артериальной. Поэтому эритроциты занимают 40% объема артериальной крови и 40,4% – венозной. Образующиеся при реакции диссоциации Н2СО3 протоны не изменяют рН, поскольку гемоглобин, будучи амфолитом, обладает значительной буферной емкостью. Кроме того, восстановленный гемоглобин является более слабой кислотой, чем оксигемоглобин, поэтому может присоединять дополнительное количество Н+. В тканях Hb ведет себя как основание:
KHbO2 ® O2 = KHb +H2CO3 ® H Н b H+
(HСО3 + H+)
II. Второй способ связывания СО2 – непосредственное его присоединение к аминогруппам белкового компонента гемоглобина за счет образования карбаминовой связи. В легких Hb выполняет функцию кислоты:
HbNH2 + CO2 ® HbNHCOO- + H+
Соединение гемоглобина с СО2 называется карбаминогемоглобином. На количество связываемого таким образом СО2 влияет оксигенация гемоглобина. Присоединение О2 к гемоглобину уменьшает количество связываемого в виде карбаминового соединения СО2.
Еще в 1892 г. Вериго было установлено, что газовая емкость по отношению к СО2 крови, содержащей восстановленный гемоглобин, значительно выше, чем в условиях полной оксигенации гемоглобина. Это явление, затем подробно изученное Холденом (1914), обусловлено тем, что, во-первых, оксигемоглобин является более сильной кислотой, чем восстановленный гемоглобин, и, следовательно, когда реакция происходит между КНbO2 и Н2СО3, равновесие наступает при образовании меньшего объема КНСО3, чем в случае, когда реакция течет между КНb и Н2СО3. Во-вторых, оксигенация гемоглобина ведет к снижению образования карбаминогемоглобина, поскольку уменьшает количество свободных NH2 групп глобина, способных связывать СО2. Следовательно, оксигенация гемоглобина влияет на способность крови связывать СО2. Это явление получило название эффекта Вериго-Холдена.
Соотношение между фракциями СО2 в крови. Каждый мл крови, протекая через ткани, захватывает примерно 2 мМоль СО2. 5-10% этого количества остается в физически растворенном виде, 10% образует карбаминовую связь с гемоглобином, 35% транспортируется в виде бикарбонатов в эритроцитах, а остальные 45% – в виде бикарбонатов натрия – в плазме. При прохождении крови через легкие СО2 выделяется из этих фракций точно в таком же соотношении.
Таким образом, дыхательная функция крови является важной составной частью функциональной системы транспорта газов и необходима для следующего этапа дыхания – газообмена между кровью и тканями.
Источник