Механизмы присоединения кислорода к гемоглобину

Кооперативное взаимодействие
Взаимовлияние протомеров олигомерного белка друг на друга называется кооперативное взаимодействие.
В легких такое взаимодействие субъединиц гемоглобина повышает его сродство к кислороду и ускоряет присоединение кислорода в 300 раз. В тканях идет обратный процесс, сродство снижается и ускорение отдачи кислорода также 300-кратное.
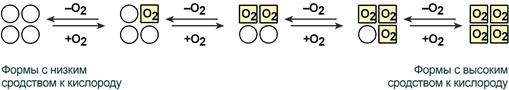
Схема кооперативного взаимодействия субъединиц гемоглобина
Объясняется такой феномен тем, что в легких при присоединении первой молекулы кислорода к железу (за счет 6-й координационной связи) атом железа втягивается в плоскость гема, кислород остается вне плоскости. Это вызывает перемещение участка белковой цепи и изменение конформации первого протомера. Такой измененный протомер влияет на другие субъединицы и облегчает связывание кислорода со второй субъединицей. Это меняет конформацию второй субъединицы, облегчая присоединение последующих молекул кислорода и изменение других протомеров.

Изменение формы субъединиц гемоглобина
при присоединении и отщеплении кислорода
Дезоксиформа гемоглобина обозначается как Т-форма, напряженная (англ. tense), она обладает существенно более низким сродством к кислороду. Оксигенированная форма, или R-форма (англ. relaxed), обладает высоким сродством к кислороду.
Изменение рН среды
Влияние рН на сродство гемоглобина к кислороду носит название эффекта Бора. При закислении среды сродство снижается, при защелачивании – повышается.
При повышении концентрации протонов (закисление среды) в тканях возрастает освобождение кислорода из оксигемоглобина. В легких после удаления угольной кислоты (в виде СО2) из крови и одновременном увеличении концентрации кислорода высвобождаются ионы Н+ из гемоглобина.
Реакция взаимодействия кислорода с гемоглобином упрощенно имеет вид:

Изменение сродства гемоглобина к кислороду в тканях и в легких при изменении концентрации ионов H+ и О2 обусловлено конформационными перестройками глобиновой части молекулы. В тканях молекула О2 отрывается от железа и ионы водорода присоединяются к остаткам гистидина (глобиновой части), образуя восстановленный гемоглобин (H-Hb) с низким сродством к кислороду. В легких поступающий в больших количествах кислород “вытесняет” ион водорода из связи с остатком гистидина гемоглобиновой молекулы.

Механизм эффекта Бора
Роль 2,3-дифосфоглицерата
2,3-Дифосфоглицерат образуется в эритроцитах из 1,3-дифосфоглицерата, промежуточного метаболита гликолиза, в реакциях, получивших название шунт Раппопорта.
Реакции шунта Раппопорта
2,3-Дифосфоглицерат располагается в центральной полости тетрамера дезоксигемоглобина и связывается с β-цепями, образуя поперечный солевой мостик между атомами кислорода 2,3-дифосфоглицерата и аминогруппами концевого валина обеих β-цепей, также аминогруппами радикалов лизина и гистидина.

Расположение 2,3-дифосфоглицерата в гемоглобине
Функция 2,3-дифосфоглицерата заключается в снижении сродства гемоглобина к кислороду, что имеет особенное значение при подъеме на высоту и при нехватке кислорода во вдыхаемом воздухе. В этих условиях связывание кислорода с гемоглобином в легких не нарушается, так как концентрация его относительно высока. Однако в тканях за счет 2,3-дифосфоглицерата отдача кислорода возрастает в 2 раза.
Связывание гемоглобина с кислородом. Обмен железаНаиболее важным свойством гемоглобиновой молекулы является ее способность свободно и обратимо связываться с кислородом. Эта способность детально изложена в нашей статье в связи с дыханием, поскольку главной функцией гемоглобина в организме является соединение его с кислородом в легких и затем освобождение этого кислорода в тканевых капиллярах, где парциальное давление кислорода гораздо ниже, чем в легких. Кислород не соединяется с двумя положительными связями железа гемоглобиновой молекулы. Вместо этого он связывается с так называемыми координационными связями атома железа. Это чрезвычайно свободная связь, поэтому соединение легко обратимо. Более того, кислород не переходит в ионную форму и переносится в виде молекулярного кислорода (составленного из двух атомов кислорода) к тканям, где легко освобождается в тканевые жидкости в форме молекулярного кислорода, а не иона кислорода. Обмен железаПоскольку железо необходимо для формирования не только гемоглобина, но и других жизненно важных элементов организма (например, миоглобина, цитохромов, цитохромоксидазы, пероксидазы, катализы), важно понять способы утилизации железа в организме. Общее количество железа в теле человека в среднем составляет 4-5 г, причем около 65% этого количества входит в состав гемоглобина. Примерно 4% железа входит в состав миоглобина, 1% находится в составе различных гем-соединений, способствующих внутриклеточному окислению, 0,1% связан с белком трансферрином в плазме крови и 15-30% накапливаются для последующего использования в основном в ретикулоэндотелиальной системе и клетках паренхимы печени главным образом в форме ферритина. Транспорт, накопление и метаболизм железа в организме схематически представлены на рисунке. После всасывания из тонкого кишечника железо немедленно связывается в плазме крови с бета-глобулином (апотрансферрином), формируя трансферрин, который затем транспортируется в плазме. Железо в трансферрине связано свободно и, следовательно, может высвобождаться в любую тканевую клетку в любой точке тела. Избыток железа в крови откладывается в основном в гепатоцитах и в меньшей степени — в ретикулоэндотелиальных клетках костного мозга. В цитоплазме клеток железо связывается главным образом с белком апоферритином, формируя ферритин. Молекулярная масса апоферритина — около 460000, и с этой большой молекулой может соединяться в кластеры радикалов железа разное количество железа; следовательно, ферритин может содержать как большое, так и небольшое количество железа. Железо, которое накапливается в виде ферритина, называют резервным железом. Меньшая часть этого резерва хранится в совершенно нерастворимой форме, называемой гемосидерином. Он в основном формируется, когда общее количество железа в теле больше, чем может связать апоферритин. Гемосидерин собирается в клетках в виде больших кластеров, которые видны под микроскопом, как большие частицы. Наоборот, частицы ферритина так малы и рассеяны, что их можно обычно увидеть в цитоплазме клеток только с помощью электронного микроскопа. Когда уровень железа в плазме падает до низких значений, некоторое его количество из запаса в форме ферритина легко выделяется в плазму и транспортируется в виде трансферрина к нуждающимся в железе областям. Уникальной особенностью молекулы трансферрина является то, что она прочно связывается с рецепторами клеточной мембраны эритробластов в костном мозге. Затем вместе со связанным железом молекула поглощается эритробластом путем эндоцитоза. Внутри клетки трансферрин освобождает железо прямо в митохондрии, где синтезируется гем. При отсутствии адекватного количества трансферрина в крови людей недостаточность транспорта железа к эритробластам может вызвать тяжелую гипохромную анемию, которая характеризуется наличием красных клеток крови, содержащих гораздо меньше гемоглобина, чем в норме. Когда эритроциты завершают свой жизненный путь и разрушаются, гемоглобин, выделившийся из клеток, поглощается моноцитарно-макрофагальными клетками. Железо при этом освобождается и накапливается преимущественно в виде ферритина, чтобы использовать ся, когда необходимо, для формирования нового гемоглобина. Ежедневная потеря железа. Мужчина выделяет примерно 0,6 мг железа ежедневно, главным образом с фекалиями. Дополнительное количество железа теряется при кровотечении. У женщин дополнительная менструальная потеря крови дает долговременную потерю железа в среднем примерно до 1,3 мг/сут. – Также рекомендуем “Всасывание железа в кишечнике. Длительность жизни эритроцитов” Оглавление темы “Эритропоэз. Белые клетки крови”: |
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 10 июля 2013;
проверки требуют 29 правок.
Эффект Вериго — Бора (синонимы — эффект Вериго, эффект Бора) — зависимость степени диссоциации оксигемоглобина от величины парциального давления углекислоты в альвеолярном воздухе и крови, при снижении которого сродство кислорода к гемоглобину повышается, что затрудняет переход кислорода из капилляров в ткани. Эффект этот был открыт независимо друг от друга Б. Ф. Вериго в 1892 году[1] и датским физиологом К. Бором в 1904 году[2].
История открытия[править | править код]
Основу разработки проблемы гипоксии заложил русский учёный-физиолог И. М. Сеченов фундаментальными работами по физиологии дыхания и газообменной функции крови. Большое значение имеют также исследования русского физиолога Б. Ф. Вериго по физиологии газообмена в лёгких и тканях. Опираясь на идеи Сеченова о сложных формах взаимодействия между диоксидом углерода и кислородом в крови (Вериго работал в лабораториях Сеченова, И. Р. Тарханова и И. И. Мечникова), он впервые установил зависимость степени диссоциации оксигемоглобина от величины парциального давления углекислоты в крови.
Биохимический механизм[править | править код]
Эффект Вериго — Бора: влияние pH на кривую насыщения гемоглобина кислородом
В клетках периферических тканей органическое топливо окисляется в митохондриях с использованием кислорода (клеточное дыхание), доставляемого гемоглобином из лёгких; при этом в качестве продуктов образуются углекислый газ, вода и другие соединения. Образование углекислого газа в тканях одновременно приводит и к повышению концентрации ионов H+ (то есть к понижению pH), поскольку при гидратации CO2 образуется H2CO3 — слабая угольная кислота, диссоциирующая на ионы H+ и бикарбонат-ионы:
H2CO3 ⇄ H+ + HCO3-.
Гемоглобин переносит значительную долю (около 20 %) общего количества CO2 и ионов H+, образующихся в тканях и поступающих в лёгкие и почки, обеспечивающих выделение этих продуктов.
За много лет до открытия этого механизма было обнаружено, что на связывание кислорода гемоглобином очень сильное влияние оказывает pH и концентрация CO2: при присоединении CO2 и ионов H+ способность гемоглобина связывать O2 снижается. Действительно, в периферических тканях с относительно низким значением pH и высокой концентрацией CO2 сродство гемоглобина к кислороду падает. И наоборот, в лёгочных капиллярах выделение CO2 и сопутствующее ему повышение pH крови приводит к увеличению сродства гемоглобина к кислороду. Это влияние величины pH и концентрации CO2 на связывание и освобождение O2 гемоглобином и называют эффектом Вериго — Бора.
Реакция связывания кислорода гемоглобином в виде
Hb + O2 ⇄ HbO2
в действительности отражает неполную картину, поскольку не учитывает дополнительные лиганды H+ и CO2.
Чтобы объяснить влияние концентрации ионов H+ на связывание кислорода, следует записать эту реакцию в иной форме:
HHb+ + O2 ⇄ HbO2 + H+,
где HHb+ — протонированная форма гемоглобина. Из этого уравнения следует, что кривая насыщения гемоглобина кислородом зависит от концентрации ионов H+. Гемоглобин связывает и O2, и ионы H+, но между этими двумя процессами существует обратная зависимость. Если парциальное давление кислорода велико (что наблюдается, например, в лёгких), то гемоглобин связывает его, освобождая при этом ионы H+. При низком парциальном давлении кислорода (что имеет место в тканях) связываться с гемоглобином будут ионы H+.
Эффективность связывания гемоглобином углекислого газа (с образованием карбаминогемоглобина, чаще именуемого карбгемоглобином[3]) находится в обратной зависимости от связывания кислорода. В тканях часть избыточного CO2 связывается с гемоглобином, сродство последнего к O2 снижается, и происходит высвобождение кислорода. В лёгких же связывается избыток O2 воздуха, тем самым сродство гемоглобина к CO2 уменьшается, и CO2 выделяется в альвеолярный воздух, способствуя лёгкому закислению крови за счёт ионов H+, возникающих при диссоциации угольной кислоты (см. выше).
Описанные зависимости делают молекулу гемоглобина великолепно приспособленной к осуществлению совместного переноса эритроцитами кислорода, углекислого газа и ионов H+[4].
См. также[править | править код]
- Эффект Холдейна (открыт Джоном Скоттом Холдейном)
Примечания[править | править код]
- ↑ Verigo B. F. Zur Frage über die Wirkung des Sauerstoff auf die Kohlensäureausscheidung in den Lungen // Archiv für die gesammte Physiologie des Menschen und der Thiere : статья. — 1892. — № 51. — С. 321—361.
- ↑ Chr. Bohr, K. Hasselbalch, and August Krogh. Concerning a Biologically Important Relationship — The Influence of the Carbon Dioxide Content of Blood on its Oxygen Binding // Skand. Arch. Physiol. : статья. — 1904. — № 16. — С. 401—412.
- ↑ Карбгемоглобин // Биологический энциклопедический словарь (рус.).
- ↑ Ленинджер А. Основы биохимии: В 3-х томах. Том 1. — М.: Мир, 1985. — С. 208—210. — 367 с.
Литература[править | править код]
- Вериго эффект // Большой медицинский словарь (рус.). — 2000.
Гемоглобин связывает и передает тканям за сутки около 600 л О2, в тканях образуется примерно 500 л СО2, который выводится из организма при участии гемоглобина.
Скорость оксигенирования и транспорт кислорода из лёгких в ткани зависит от изменений сродства гемоглобина к О2.
Присоединением О2 к одному из протомеров гемоглобина в лёгких при высоком рО2, (100 мм рт. ст.) вызывает конформационные изменения во всей олигомерной структуре гемоглобина и облегчает присоединение последующих молекул О2. Это явление носит название кооперативных изменений гемоглобина или кооперативного связывания О2, благодаря которому гемоглобин максимально связывает О2 в легких и легко отдает О2 в периферических тканях при более низких значениях рО2 (рис кривая насыщения миоглобина и гемоглобина).
Давление в капиллярах Давление в лёгочных
работающей мышцы а л ь в е о л а х
Рис.Кривые насыщения (диссоциации) миоглобина и гемоглобина кислородом.
Кривая насыщения для гемоглобина имеет сигмоидную форму (S-образную). Это указывает на кооперативное связывание О2 (в легких при высоком рО2) и освобождении О2 (в тканевых капиллярах при более низком рО2).
Эффект Бора
Гемоглобин не только переносит О2 от легких к периферическим тканям, но и ускоряет транспорт СО2 и протонов от тканей к легким.
Эффект Бора – это влияние концентрации СО2 и Н+ на сродство гемоглобина к О2.
Увеличение концентрации протонов и СО2 снижает сродство О2 к гемоглобину и усиливает транспорт О2 в ткани. Разные органы потребляют неодинаковые количества кислорода. Наиболее активно извлекают кислород из крови работающие мышцы (до 85%).
Если гемоглобин насыщен кислородом почти на 100%, то все молекулы гемоглобина находятся в R-форме, Нb(О2)4. Такое состояние гемоглобина возникает при высоком парциальном О2в капиллярах легких (100 мм рт. ст.) и при высоком сродстве Нb к О2. В капиллярах мышц и других тканей более низкое парциальное давление О2 , в этих условиях от гемоглобина могут отщепляться молекулы О2 и поступать в ткани. Например, при парциальном давлении кислорода в капиллярах 40 мм рт. ст. гемоглобин имеет насыщение ≈ 65%.
Далее действуют другие факторы, приводящие к снижению сродства гемоглобина к О2, тем самым увеличивая доставку кислорода в ткани:
а) гемоглобин протонируется, связывая три пары Н+ в пептидных цепях гемоглобина, вновь формируется Т-структура;
б) гемоглобин обратимо присоединяет часть СО2 (до 15%) к концевым аминогруппам пептидных цепей, образуется карбогемоглобин: R-NH-COO- +Н+, где R-белковая цепь гемоглобина. Это также снижает сродство гемоглобина к О2. Большая часть СО2 транспортируется кровью в виде бикарбоната НСО3 -.
Таким образом, диоксид углерода, образовавшийся в тканях реагирует с водой, образуя угольную кислоту, которая диссоциирует на бикарбонат- ион и Н+. Синтез углекислоты, как и её разложение на СО2 и Н2О происходит при участии фермента карбоангидразы. В периферических тканях гемоглобин отдает О2, сам протонируется и связывает часть СО2, его R-форма переходит в Т-форму, сродство к О2 резко снижается. В легких происходят обратные процессы.
Влияние 2,3-БФГ на сродство гемоглобина к кислороду
2,3-биcфосфоглицерат (2,3-дифосфоглицерат) – образуется в эритроцитах в ходе анаэробного гликолиза из промежуточного соединения 1,3-биcфосфоглицерата. 2,3-БФГ является аллостерическим регулятором гемоглобина (от греч. «аллос» – другой), он соединяется ионными связями с дезоксигемоглобином не в области гема, а в другом участке, т. е. аллостерическом центре, содержащим положительно заряженные радикалы аминокислот Лиз 82 и Гис 143 β-цепей.
Структура 2,3-бифосфоглицерата
Образовавшиеся дополнительные солевые мостики (или ионные связи) делают структуру дезоксигемоглобина ещё более жесткой и поэтому сродство Нb к О2 снижается. Концентрация 2,3-БФГ в крови близка к концентрации гемоглобина и одинакова в артериальной и венозной крови. 2,3-БФГ реагирует только с дезоксигемоглобином, т. е. Т-формой, т. к. он имеет доступную центральную полость с центром связывания 2,3-БФГ. В оксигемоглобине (R-форма) эта полость закрыта.
К Т (т. е. дезокси-)-структуре. Ясно, что выяснение столь тонкой вещи, как молекулярный механизм кооперативного связывания кислорода гемоглобином, представляет собой весьма сложную задачу. [c.312]
Часто вместо уравнения (7.69) связывание кислорода гемоглобином описывают уравнением Хилла [c.233]
Гемы входят в состав гемоглобина, выполняющего в организме функцию переносчика кислорода. Активным центром в процессе связывания кислорода является атом железа (II) гема. Процесс присоединения кислорода обратим в легких, где парциальное давление кислорода высокое,, молекула Од присоединяется к атому железа, а в тканях, где парциальное давление кислорода низкое, кислород освобождается. [c.587]
Молекула гемоглобина человека, подобно гемоглобину других млекопитающих, состоит из четырех полипептидных цепей (каждая из которых содержит одну гем-группу) и способна обратимо присоединять четыре молекулы кислорода. Уже много лет назад было показано, что равновесное связывание кислорода гемоглобином описывается S-образной кривой, приведенной на рис. 15.12, которая отличается от аналогичной кривой для миоглобина. Для миоглобина, содержащего одну гем-группу в молекуле, следует ожидать кривую равновесия, отвечающую реакции [c.440]
Рассмотрим теперь вопрос о природе кооперативного связывания кислорода с тетрамерной ( 2 2) молекулой гемоглобина (разд. Г.8) и физиологическое значение этого процесса [65]. Полипептидная цепь [c.304]
Далее, путем модификации остатка пропионовой кислоты в боковой цепи порфиринового кольца был введен второй имидазольный лиганд, соответствующий проксимальному гистидину природных переносчиков кислорода. Интересно, что все структурные элементы активного центра миоглобина или гемоглобина, которые существенны для связывания кислорода, присутствуют [c.368]
Связывание кислорода гемоглобином можно описать математически следующим набором равновесий, где НЬ обозначает молекулу гемоглобина, которая может присоединять четыре молекулы кислорода [c.232]
Изменение конформации полипептидных цепей гемоглобина при связывании кислорода — пример так называемой аллостерии. Известны аллостерические формы и у других белков, преимущественно у фермен- [c.443]
Связывание кислорода гемоглобином демонстрирует особые свойства, которые может проявлять белок в реакциях присоединения. В отличие от большинства реакций с участием малых молекул сродство белка к лиганду может возрастать по мере присоединения все новых молекул лиганда. [c.211]
Приведенные выше биохимические равновесия включали небольшие молекулы, однако во многих таких равновесиях участвуют макромолекулы, например белки и нуклеиновые кислоты. В качестве примера рассмотрим связывание кислорода гемоглобином. [c.231]
СВЯЗЫВАНИЕ КИСЛОРОДА МИОГЛОБИНОМ И ГЕМОГЛОБИНОМ [c.231]
Кооперативный характер связывания ферментов с субстратами имеет, пожалуй, такое же большое физиологическое значение, как и кооперативное связывание гемоглобина с кислородом, которое обеспечивает более эффективное высвобождение связанного кислорода в тканях (гл. 4, разд. Д, 5). Кооперативность связывания субстрата отсутствует в том случае, когда благодаря избытку активатора фермент переходит в состояние R (В), при котором связывающие центры ведут себя независимо. В то же время связывание активатора должно характеризоваться сильно выраженной кооперативностью, т. е. скорость реакции должна изменяться при изменении концентрации активатора сильнее, чем в случае гиперболической активации. Аналогичным образом кооперативное связывание ингибитора обеспечивает более быстрое выключение фермента при увеличении концентрации ингибитора. По-видимому, эволюция олигомерных ферментов (по крайней мере отчасти) обусловлена большей эффективностью механизмов регуляции, в основе которых лежит кооперативное связывание эффекторов. [c.39]
Гемоглобин по своему строению гомологичен миоглобину и практически представляет собой тетрамер миоглобина — оба белка действуют взаимосвязанно в биологических системах. Такая кооперация в действии, помимо прочего, требует от гемоглобина высокого сродства к кислороду при его высоком парциальном давлении и низкого сродства при недостатке кислорода. Рис. 3-39 графически демонстрирует связывание кислорода гемоглобином и миоглобином. Форма кривой сигмоидная для гемоглобина и гиперболическая с крутым подъемом для миоглобина. [c.415]
Читателю предоставляется интересная возможность проанализировать, можно ли на основании этого уравнения предсказать слабое кооперативное связывание кислорода гемоглобином миноговых. [c.302]
Для них характерна большая зависимость между конформацией белковой молекулы и каталитической активностью. Их действие не подчиняется кинетике Михаэлиса-Ментен и описывается другими уравнениями, а графически зависимость скорости от концентрации субстрата имеет сигмоидный характер. Для аллостерических ферментов характерно проявление кооперативного эффекта, когда связывание одной молекулы субстрата усиливает способность присоединять следующую молекулу (активирующий кооперативный эффект), что видно на примере гемоглобина при связывании кислорода с одной субъединицей усиливается дальнейшее взаимодействие его с другими субъединицами. Тот факт, что аллостерический эффект проявляется часто на первой стадии процесса, объясняет выработанное в ходе эволюции экономное расходование веществ на последующих стадиях реакции. [c.35]
Ранее отмечалось, что связывание кислорода атомом железа гема переводит последний из непланарной в планарную конфигурацию. Этот процесс запускает другие конформационные изменения, которые приводят к такому кооперативному взаимодействию между субъединицами гемоглобина, что, как только некоторое количество кислорода связывается, связывание последующих молекул облегчается. Следует теперь рассмотреть этот процесс в деталях. Предпоследним аминокислотным остатком как в а-, так и [c.558]
Рнс. 5.8. Связывание кислорода гемом в гемоглобине. [c.170]
Кооперативность связывания кислорода с гемоглобином была открыта очень давно, и, несмотря на это, важность данного явления недооценивали. Оно вновь привлекло к себе широкое внимание в 1965 г., когда Moho, Уаймен и Шанжё [33] описали его математически. Поскольку для многих случаев предложенная авторами модель является сильным упрощением, ниже мы остановимся на более общем подходе к этому вопросу, разработанном Кошландом [60—62]. [c.297]
Детали механизма связывания кислорода гемоглобином сложны, но хорошо изучены. Будучи чрезвычайно важным с физиологической точки зрения, этот процесс служит прекрасной иллюстрацией аллостерических взаимодействий и регуляции. Характерные особенности связывания кислорода гемоглобином могут быть суммированы следующим образом [c.170]
Наконец, следует напомнить, что железо, связанное с порфи-рииом (гем), находится в ферросостоянии. Процесс связывания кислорода гемоглобином обратим, причем молекула кислорода и атом лелеза находятся в стехиометрическом соотношении 1 1 и не происходит окисления Ре(П) до Ре(П1). Исследованию такого обратимого связывания молекулярного кислорода с Ре(П) в геме уделено очень большое внимание. Способность гема обратимо связывать кислород, проявляется при его включении в большую белковую структуру. Одиако если гем извлечь из белка и поместить в раствор при комнатной температуре, молекулярный кислород необратимо окисляет железо до феррисостояния Ре(П1). [c.361]
В гемоглобине атом Ре(П) прикрыт (экранирован) в плоскости молекулы гема атомами макроцикла, а сверху и снизу — остатками имвдазола (Im) аминокислотного фрагмента гистидина (His), находящегося в составе молекулы белка. Один из имндазолов Irai образует донорно-акцепторную связь с железом, а второй Imi находится на значительном удалении от железа и не координирован с ним. Второй имидазол бережет шестое координационное место во внутренней координационной сфере железа (П) для молекулы О2, не допуская к нему никаких молекул, даже Н2О. Сам химизм обратимого связывания кислорода воздуха прост [c.745]
Если в случае дезоксигемо-глобина никакой заметной диссоциации тетрамера на субъединицы не наблюдается, то оксиге-моглобин слабо диссоциирует на ар-димеры (/С = 2-10- ). Вопросу связывания гемоглобинов с кислородом посвящено огромное [c.306]
Рассмотренные выще механизмы способны описывать многие сложные эффекты, и кинетическое уравнение может иметь очень сложную форму. Но в общем случае концентрация [ЕЗ] не может возрастать быстрее, чем растет [3]. Однако при некоторых экспериментальных условиях субстраты или ингибиторы оказывают большее влияние на концентрацию комплекса. Другими словами, получаются 3-образные кривые типа кривой связывания кислорода гемоглобином (разд. 7.13). В особенности это относится к ферментам, играющим важную роль в регулировании обмена веществ. Подобные кооперативные эффекты встречаются в случае ферментов с несколькими активными центрами, поскольку кооперативный эффект подразумевает возрастание сродства второго активного центра к субстрату, когда первый центр занят. Как и в случае гемоглобина, взаимодействия такого типа сопровождаются структурными изменениями. Согласно модели Моно — Шанжо — Ваймана, фермент с несколькими активными центрами может находиться по крайней мере в двух состояниях. Это, вероятно, слишком упрощенная картина, но два является минимальным числом состояний, необходимым для объяснения наблюдаемых эффектов. Предполагается, что в обоих состояниях конформации всех субъединиц одинаковы. Воздействующая на систему молекула (эффектор), которая может быть молекулой субстрата, смещает равновесие в сторону одного или другого из этих двух состояний. Если эффектор смещает равновесие в направлении увеличения скорости реакции, то такой эффектор называется активатором. Если же его действие приводит к снижению скорости реакции, то он называется ингибитором. Как и в случае гемоглобина, воздействие усиливается тем, что одна молекула эффектора оказывает влияние на несколько каталити-21 [c.323]
Это означает, что при значениях р(02), обычно характерных для плаценты, кислород будет переходить с НЬА на НЬР с выгодой для плода. Кооперативность связывания кислорода гемоглобином А выражена настолько явно, что, если молекула НЬА уже связала три молекулы кислорода, то вероятность связывания четвертой молекулы в 70 раз превосходит вероятность приобретения дезоксиге-моглобином одной молекулы кислорода. [c.558]
Рассмотрим случай, когда константа Къв очень мала и Ва легко диссоциирует на мономеры. Тогда присоединение X приведет к диссоциации димера. Хорошо известным примером белка такого рода может служить гемоглобин миноговых, который представляет собой димер и после связывания кислорода диссоциирует на мономеры [64]. В этом случае уравнение (4-49) сводится к такому виду [c.302]
Что же общего между всеми гемоглобинами Прежде всего для них характерен один и тот же способ укладки полипептидных цепей вокруг идентичных для всех гемоглобинов (или очень сходных) гемогрупп. Однако наиболее поразительным является тот факт, что, несмотря на четко выраженное единообразие общей структуры всех гемоглобинов, имеется всего девять инвариантных аминокислотных остатков и один почти инвариантный. Эти десять остатков заключены на рис. 4-17 в прямоугольные рамки. Два глицина (или аланина) в положениях В-6 и Е-8 инвариантны потому, что тесный контакт между спиралями В и Е не позволяет находиться в этих положениях аминокислотным остаткам большего размера. Пролин С-2 обеспечивает изгиб молекулы. Четыре других инвариантных остатка непосредственно связаны с гемогруппой. Два из них. His Е-7 и His F-8, являются гем-связанными гистидинами. Девятый остаток. Туг НС-2, о котором уже шла речь в разд. 5.а, играет основную роль в кооперативном связывании кислорода. И только Lys Н-9 расположен с наружной стороны молекулы. Причины, по которым этот остаток инвариантен, не ясны [80]. [c.314]
Было сделано необычайно интересное открытие, заключающееся в том, что координация гемового железа с гистидином, по всей видимости, лежит в основе кооперативности при связывании гемоглобина с кислородом [9, 10]. Радиус высокоспинового железа как в ферри-, так и в ферросостоянии столъ велик, что железо не помещается в центре порфиринового кольца и Смещается в сторону координационно связан- ной с ним имидазольной группы на расстояние, составляющее для Ре(II) 0,06 нм. Таким образом, в дезоксигемоглобине и железо, и имидазольная группа находятся от плоскости кольца дальше, чем в оксиге-моглобине. В последнем железо находится в центре порфиринового кольца, так как переход в низкоспиновое состояние сопровождается уменьшением ионного радиуса [9, И]. Изменение конформации белка, индуцированное небольшим смещением иона железа, уже было описано (гл. 4, разд. Д, 5). Однако истинная природа пускового механизма , приводящего к этим изменениям, пока в точности не ясна. С некоторым атриближением эти изменения можно рассматривать как чисто механи- [c.368]
Механизм действия сиГнильной кислоты заклшается в необратимом ингибировании железосодержащих дыхательных ферментов. Вследствие сильного сродства цианид- аниона к иону цитохромоксидазы ак тивность этого фермента уменьшается, в результате чего прекращаются процессы клеточного окисления, управляемые атим ферментом, которые составляют свыше 90% все дыхательной деятельности клет-I ки Подобно гемоглобину, функцией цитохромоксидазы является об– ратимое связывание кислорода и двуокиси углерода. [c.3]
Биологическое значение маскировки а-аминогрупп недостаточно ясно возможно, она защищает белок от атаки аминопептидаз или способствует закреплению N-концевой части полипептида в аполярном окружении либо на молекуле рецептора, либо внутри самого белка, чтобы препятствовать его контакту с раствором. Это предположение не относится к метилированию а-аминогруппы, обнаруженному в рибосомных белках, выделенных из Es heri hia oli [135], поскольку метилирование не элиминирует заряд. Физиологическая роль ацетилирования а-аминогруппы совершенно ясна в случае некоторых гемоглобинов рыб такая модификация помогает сохранять способность к связыванию кислорода независимо от рН-среды, что предотвращает выделение избыточного кислорода в плавательный пузырь [136] (разд. 10.3). [c.72]
Дана зависимость насыщения гемоглобина (НЬ) кислородом при pH 7,2 от концентрации свободного кислорода. Концентрации 0 в капиллярах легких (125 мкМ) н в капиллярах тканей, потребляющих (50 мкМ), зафиксированы в узких пределах. Кривая а в отсутствие дифосфоглицерата (ДФГ) гемоглобин насыщается О в легких, но не может доставлять его к тканям. Кривая б прн физиологическом уровне ДФГ (4.5 мМ. приблизительно 30% Оа. поглощенного легкими, высвобождается в тканях (стрелка 1). Кривые бив поскольку гемоглобин плода (кривая в) имеет более низкое сродство к ДФГ. чем материнский гемоглобин, освобожденный из материнской крови молекулярный кислород может захватываться гемоглобином плода (стрелка [[I). Кривая г высокая концентрация ДФГ (8 мМ) приводит к повышенному снабжению тканей кислородом (стре.жи I и [[). Кривая д при отсутствии кооператнвиостн между субъединицами гемоглобина от легких к тканям транспортировалось бы меньше Оз. При построении гипотетической кривой связывания (5) для комплекса НЬОз принята константа диссоциации 38 мкМ. [c.258]
Кривая связывания кислорода мио-глобином представляет собой ректан-гулярную гиперболу, в то время как для гемоглобина она сигмоидальна ввиду аллостерического характера связывания (рис. 24.2.1). За счет этого достигается преимущество в биологической функции, поскольку присоединение и освобождение кислорода происходит в небольшом интервале (Ог). [c.557]
Кооперативное связывание кислорода. Когда молекула гемоглобина диссоциирована на составляющие ее полипептидные цепи, поведение последних очень сходно с поведением миоглобина их кривые освобождения кислорода имеют гиперболиче- [c.171]
Кривая связывания кислорода гемогл