Обмен гемоглобина образование билирубина


В норме обмен билирубина включает 3 этапа: образование из гемоглобина эритроцитов, превращение токсичного непрямого в водорастворимый прямой, выведение последнего с желчью через кишечник. Если на первом этапе усиленно разрушаются эритроциты (гемолитическая анемия, сепсис, малярия), то возрастает содержание непрямого билирубина, а надпеченочная желтуха характеризуется темным (почти черным) калом и лимонным цветом кожи.
При болезнях печени (гепатит, цирроз) нарушения связывания пигмента происходят на втором этапе, повышается прямой и непрямой билирубин. Внутрипеченочная желтуха протекает с увеличением печени, возможно ухудшение сознания, кожа с оранжевым оттенком, темная моча.
Для болезней с патологией обмена на третьем этапе типично затруднение оттока желчи (камни желчных путей, опухоль). Из-за этого растет прямой билирубин, а подпеченочная желтуха имеет признаки: зеленоватый цвет кожи, светлый кал и моча цвета темного пива.
Что означает обмен билирубина, по какой схеме он происходит
Обмен билирубина – это цикл из последовательных этапов биологических реакций, который можно представить в виде схемы из 3 основных этапов:
- Распад эритроцитов и образование билирубина.
- Поступление в печень и обезвреживание.
- Выведение пигмента из организма.
Распад эритроцитов и биохимия обмена в крови
Первый этап биохимии пигментного обмена – это цикл реакций от распада клеток крови до образования билирубина. Главный источник желчного пигмента – это гемоглобин эритроцитов (80%), немного (20%) образуется из гемовых белков клеток-предшественников (например, незрелых ретикулоцитов или эритробластов).
Местом превращений является костный мозг, селезенка и печень. Кровяная клетка разрушается через 3 месяца жизни и освободившийся гемоглобин распадается на:
- глобин (белок, он превращается в аминокислоты, идущие потом на синтез ферментов);
- гемосидерин с железом, которое окисляется и запасается в виде ферритина;
- гематоидин переходит в биливердин, а затем в билирубин.
Образовавшийся пигмент соединяется с мелкими белками крови (альбуминами) и с кровотоком поступает в печень.
Обезвреживание в печени
Печеночный этап обмена включает такие стадии:
- Образование непрямого билирубина.
- Превращение непрямого в прямой.
- Выделение прямого в желчные пути.
Билирубиново-альбуминовый комплекс вначале поступает в синусы печени, где от него отщепляется белковая часть и в результате появляется токсичный для организма непрямой (несвязанный) билирубин. Он опасен для организма, так как хорошо растворим в жирах, может проникать через клеточные мембраны и накапливаться в тканях, в том числе и нейронах мозга.
Непрямой билирубин проникает в клетки печени, в которых происходит очень важное обезвреживание пигмента путем соединения с глюкуроновой кислотой. В результате теряется способность к его растворению в жирах и пигмент приобретает гидрофильность, то есть растворимость в воде. Эта особенность помогает ему:
- поступать в желчь;
- частично фильтроваться в почках (окрашивает мочу в желтый цвет);
- быстро вступать в соединение с реактивами при исследовании крови, поэтому он и называется прямым, так как сразу реагирует (прямая и быстрая реакция).
Прямой билирубин из клеток печени переходит в желчные протоки. На этой стадии процесс контролируется гипофизом головного мозга и гормонами щитовидной железы. Билирубин в желчи соединяется с холестерином, жирными кислотами, белками. Содержимое протоков печени поступает в желчный пузырь, потом при приеме пищи оно передвигается в кишечник для улучшения переваривания жиров.
Выведение из организма
В кишечнике из билирубина под действием бактерий образуется уробилиноген и стеркобилиноген. Из последнего получается стеркобилин, окрашивающий кал в коричневый цвет. Часть уробилиногена поступает обратно в кровь из кишечной стенки и в печени полностью перерабатывается. Небольшое количество пигментов выводится с мочой.
Смотрите в этом видео об обмене билирубина:
Механизм образования билирубина и его особенности у детей
У новорожденных есть отличия механизма образования и обмена билирубина, они связаны с особенностями клеточного состава крови и незрелостью ферментов, участвующих в превращении и выведении пигмента. Это проявляется в виде физиологической (то есть без заболеваний) желтухи. Основные причины:
- недостаточность ферментной системы печени, которые участвуют в превращении непрямого билирубина в прямой (у недоношенных может продолжаться до 3 месяцев);
- много образуется в костном мозге клеток-предшественников;
- эритроциты быстрее разрушаются, чем у взрослых.
Схема патогенеза гемолитической желтухи у новорожденных
Желтуха без нарушений состава крови и работы печени не ухудшает самочувствия ребенка и проходит самостоятельно.
Метаболизм билирубина при заболеваниях
Нарушения метаболизма билирубина могут возникать на разных этапах:
- разрушение эритроцитов – гемолитические анемии, малярия, талассемия, аутоиммунные анемии;
- отклонения захвата билирубина из крови и превращения непрямого в прямой – вирусные, токсические гепатиты, цирроз печени, жировое перерождение, опухоль;
- препятствия на пути выведения в желчные пути или поступления в кишечник – камни, сужения протоков, сдавление опухолью или увеличенной головкой поджелудочной железы.
В зависимости от уровня поражения выделено три типа желтухи:
- надпеченочная (при заболеваниях крови);
- печеночная (разрушение клеток печени);
- подпеченочная (затруднение оттока желчи).
Почему так важно, чтобы был обмен билирубина в норме
Если на каком-либо этапе обмен билирубина отклоняется от нормы, то возникает:
- пожелтение кожи и слизистых оболочек из-за накопления пигмента;
- замедляется образование энергии и белковых молекул (слабость, низкая работоспособность);
- затруднения переваривания пищи (вздутие, нерегулярный стул, самоотравление организма);
- боль в области печени (правое подреберье);
- ухудшение работы печеночных клеток.
Самое опасное состояние связано с накоплением токсичного для организма непрямого билирубина. Он обладает способностью разрушать скопления клеток головного мозга (ядер) у новорожденных. Развивается энцефалопатия с ухудшением сознания и неврологическими отклонениями (судороги, нарушения дыхания, слуха), она в тяжелом случае сопровождается коматозным состоянием и летальным исходом.
Для взрослых пациентов характерно замедление выведения холестерина, что увеличивает риск инфаркта, инсульта.
Причины нарушения обмена билирубина
К главным причинам нарушения обмена билирубина относятся:
- повышение непрямого из-за усиленного гемолиза (гемолитическая анемия, серповидно-клеточная анемия, талассемия, аутоиммунные болезни) и врожденных болезней с неполноценными ферментами (синдром Жильбера);
- повышения прямого, их подразделяют на печеночные, внутрипеченочных протоков и внепеченочные факторы.
Патогенез синдрома Жильбера
К печеночным заболеваниям (гепатитам) относятся: вирусный, бактериальный, алкогольный, лекарственный, токсический. Причинами застоя желчи внутри печени бывают:
- хронический гепатит в стадии обострения;
- желтуха у беременных;
- цирроз печени;
- наследственные болезни: Дабина-Джонсона, Ротора;
- раковая опухоль печеночной ткани.
Внепеченочные факторы нарушения обмена билирубина включают:
- воспаление поджелудочной железы – острое, обострение хронического;
- патологии желчных протоков – сдавление опухолью извне, рубцовые сужения, воспаление, камни;
- паразитарные заболевания – эхинококкоз;
- поражение двенадцатиперстной кишки (воспаление, опухоль);
- изменения лимфоузлов (лимфогранулематоз).
Диагностика состояния
Для уточнения диагноза, который вызвал нарушения обмена билирубина, проводят комплексное обследование:
- анализ крови общий (гемоглобин, эритроциты, их число и форма) и биохимический с печеночными тестами (билирубин и его прямая, непрямая фракции, ферменты АСТ, АЛТ, щелочная фосфатаза), ферритин, трансферрин;
- иммунология – антитела к вирусам и аутоиммунные;
- анализ мочи на билирубин, уробилин;
- анализ кала на скрытую кровь;
- УЗИ печени, селезенки;
- магнитно-резонансная томография;
- биопсия печени;
- заполнение контрастом желчных путей (холангиография).
Как справиться с проблемами
После постановки диагноза для лечения назначают:
- диету с частым, дробным питанием, запретом жирной, жареной и острой пищи, копчений, алкоголя;
- фототерапия (лечение светом);
- переливание крови или плазмы.
Фототерапия
При затруднении оттока желчи и отсутствии показаний к операции показан Урсофальк, Аллохол, Холензим, желчегонные чаи с календулой, бессмертником, тысячелистником, энтеросорбенты (Полисорб). При вирусном гепатите необходимы противовирусные средства, предупредить гемолиз помогают гормоны, цитостатики.
Если обнаружены камни в желчном пузыре, то нужна операция для его удаления. При тяжелом течении гемолитической анемии иногда приходится удалить селезенку. Опухоли, сдавливающие протоки, также подлежат хирургическому лечению.
Рекомендуем прочитать о показателях нормы билирубина в крови у женщин. Из статьи вы узнаете о причинах для проведения анализа, симптомах отклонений, таблице норм билирубина по возрасту, при беременности.
А здесь подробнее о показателях билирубина в крови у мужчин.
Нарушения обмена билирубина проявляются желтухой, она бывает связана с разрушением эритроцитов, заболеваниями печени, затруднением оттока желчи. Для постановки диагноза учитывают симптомы и данные анализов на общий, прямой, непрямой билирубин, результаты дополнительного обследования.
Источник
За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов.
Эритроциты в норме живут 90-120 дней, после чего лизируются в клетках ретикулоэндотелиальной системы – макрофагах селезенки (главным образом), купферовских клетках печени и макрофагах костного мозга. При разрушении эритроцитов в кровеносном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и также переносится в клетки РЭС селезенки, печени и костного мозга.
Синтез билирубина
В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина. Высвобождаемое железо может либо запасаться в клетке в комплексе с ферритином, либо выделяться наружу и связываться с трансферрином.
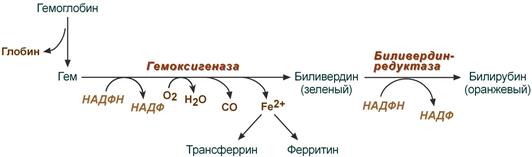
Реакции распада гемоглобина и образования билирубина
Билирубин – токсичное, жирорастворимое вещество, способное разобщать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
Строение билирубина
Выведение билирубина
Из клеток ретикуло-эндотелиальной системы билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.
Этапы метаболизма билирубина в организме
Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (транспортный белок органических анионов) или по механизму флип-флоп. Далее при участии цитозольного связывающего белка лигандина (Y-протеин) билирубин транспортируется в ЭПР, где протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию конъюгации могут вступать сульфаты, фосфаты, глюкозиды.
Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин. Одновременно, даже в норме (особенно у взрослых), некоторое количество билирубин-глюкуронидов может попадать из желчи в кровь по межклеточным щелям.
Таким образом, в плазме крови обычно присутствуют две формы билирубина: свободный (непрямой), попадающий сюда из клеток РЭС (80% и более всего количества), и связанный (прямой), попадающий из желчных протоков (в норме не более 20%).
Термины “связанный“, “конъюгированный“, “свободный“, “несвязанный” отражают взаимодействие билирубина и глюкуроновой кислоты (но не билирубина и альбумина!).
Термины “прямой” и “непрямой” введены, исходя из возможности химической реакции билирубина с диазореактивом Эрлиха. Связанный билирубин реагирует с реактивом напрямую, без добавления дополнительных реагентов, т.к. является водорастворимым. Несвязанный (жирорастворимый) билирубин требует добавочных реактивов, реагирует не прямо.
Превращение в кишечнике
В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть уробилиногена всасывается и с кровью портальной вены попадает в печень, где либо распадается до моно-, ди- и трипирролов, либо окисляется до билирубина и снова экскретируется. При этом при здоровой печени в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена. Далее
- малая часть стеркобилиногена может всасываться и катаболизировать в печени, подобно уробилиногену,
- незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и в мочу. После окисления на воздухе из стеркобилиногена образуется стеркобилин мочи,
- однако основное количество стеркобилиногена достигает нижних отделов толстого кишечника и выделяется. В прямой кишке и на воздухе стеркобилиноген окисляется в стеркобилин, окрашивая кал,
- аналогично уробилиноген, появляющийся в моче при патологии печени, окисляется в уробилин.
Очень часто стеркобилиноген, содержащийся в нормальной моче, называют уробилиногеном. И в клинической практике обычно не проводят различий между стеркобилиногеном и уробилиногеном мочи, их рассматривают как один пигмент – урохромы (уробилиноиды), что может создавать некоторую путаницу при оценке результатов анализа.
Источник
Билирубин представляет собой конечный продукт распада гема. Основная часть (80-85%) билирубина образуется из гемоглобина и лишь небольшая часть – из других гемсодержащих белков, например цитохрома Р450. Образование билирубина происходит в клетках ретикулоэндотелиальной системы. Ежедневно образуется около 300 мг билирубина.
Преобразование гема в билирубин происходит с участием микросомального фермента гемоксигеназы, для работы которого требуются кислород и НАДФН. Расщепление порфиринового кольца происходит селективно в области метановой группы в положении а. Атом углерода, входящий в состав a-метанового мостика, окисляется до моноксида углерода, и вместо мостика образуются 2 двойные связи с молекулами кислорода, поступающими извне. Образующийся в результате этого линейный тетрапиррол по структуре является IX-aльфа-биливердином. Далее он преобразуется биливердинредуктазой, цитозольным ферментом, в IX-альфа-билирубин. Линейный тетрапиррол такой структуры должен растворяться в воде, в то время как билирубин является жирорастворимым веществом. Растворимость в липидах определяется структурой IX-альфа-билирубина – наличием 6 стабильных внутримолекулярных водородных связей. Эти связи можно разрушить спиртом в диазореакции (Ван ден Берга), в которой неконъюгированный (непрямой) билирубин превращается в конъюгированный (прямой). In vivo стабильные водородные связи разрушаются этерификацией с помощью глюкуроновой кислоты.
Около 20% циркулирующего билирубина образуется не из гема зрелых эритроцитов, а из других источников. Небольшое количество поступает из незрелых клеток селезёнки и костного мозга. При гемолизе это количество увеличивается. Остальной билирубин образуется в печени из гемсодержащих белков, например миоглобина, цитохромов, и из других неустановленных источников. Эта фракция увеличивается при пернициозной анемии, эритропоэтической уропорфирин и при синдроме Криглера-Найяра.
Транспорт и конъюгация билирубина в печени
Неконъюгированный билирубин в плазме прочно связан с альбумином. Только очень небольшая часть билирубина способна подвергаться диализу, однако под влиянием веществ, конкурирующих с билирубином за связывание с альбумином (например, жирных кислот или органических анионов), она может увеличиваться. Это имеет важное значение у новорождённых, у которых ряд лекарств (например, сульфаниламиды и салицилаты) может облегчать диффузию билирубина в головной мозг и таким образом способствовать развитию ядерной желтухи.
Печенью выделяются многие органические анионы, в том числе жирные кислоты, жёлчные кислоты и другие компоненты жёлчи, не относящиеся к жёлчным кислотам, такие как билирубин (несмотря на его прочную связь с альбумином). Исследования показали, что билирубин отделяется от альбумина в синусоидах, диффундирует через слой воды на поверхности гепатоцита. Высказанные ранее предположения о наличии рецепторов альбумина не подтвердились. Перенос билирубина через плазматическую мембрану внутрь гепатоцита осуществляется с помощью транспортных белков, например транспортного белка органических анионов, и/или по механизму «флип-флоп». Захват билирубина высокоэффективен благодаря его быстрому метаболизму в печени в реакции глюкуронидизации и выделению в жёлчь, а также вследствие наличия в цитозоле связывающих белков, таких как лигандины (глутатион-8-трансфераза).
Неконъюгированный билирубин представляет собой неполярное (жирорастворимое) вещество. В реакции конъюгации он превращается в полярное (водорастворимое вещество) и может благодаря этому выделяться в жёлчь. Эта реакция протекает с помощью микросомального фермента уридиндифосфатглюкуронилтрансферазы (УДФГТ), превращающего неконъюгированный билирубин в конъюгированный моно- и диглюкуронид билирубина. УДФГТ является одной из нескольких изоформ фермента, обеспечивающих конъюгацию эндогенных метаболитов, гормонов и нейротрансмиттеров.
Ген УДФГТ билирубина находится на 2-й паре хромосом. Структура гена сложная. У всех изоформ УДФГТ постоянными компонентами являются экзоны 2-5 на 3′-конце ДНК гена. Для экспрессии гена необходимо вовлечение одного из нескольких первых экзонов. Так, для образования изоферментов билирубин-УДФГТ 1*1 и 1*2 необходимо вовлечение соответственно экзонов 1А и ID. Изофермент 1*1 участвует в конъюгации практически всего билирубина, а изофермент 1*2 почти или вовсе не участвует в этом. Другие экзоны (IF и 1G) кодируют изоформы фенол-УДФГТ. Таким образом, выбор одной из последовательностей экзона 1 определяет субстратную специфичность и свойства ферментов.
Дальнейшая экспрессия УДФГТ 1*1 зависит также от промоторного участка на 5′-конце, связанного с каждым из первых экзонов. Промоторный участок содержит последовательность ТАТАА.
Детали строения гена важны для понимания патогенеза неконъюгированной гипербилирубинемии (синдромы Жильбера и Криглера-Найяра), когда в печени содержание ферментов, ответственных за конъюгацию, снижено или они отсутствуют.
Активность УДФГТ при печёночно-клеточной желтухе поддерживается на достаточном уровне, а при холестазе даже увеличивается. У новорождённых активность УДФГТ низкая.
У человека в жёлчи билирубин представлен в основном диглюкуронидом. Превращение билирубина в моноглюкуронид, а также в диглюкуронид происходит в одной и той же микросомальной системе глюкуронилтрансферазы. При перегрузке билирубином, например при гемолизе, образуется преимущественно моноглюкуронид, а при уменьшении поступления билирубина или при индукции фермента возрастает содержание диглюкуронида.
Наиболее важное значение имеет конъюгация с глюкуроновой кислотой, однако небольшое количество билирубина конъюгируется с сульфатами, ксилозой и глюкозой; при холестазе эти процессы усиливаются.
В поздних стадиях холестатической или печёночно-клеточной желтухи, несмотря на высокое содержание в плазме, билирубин в моче не выявляется. Очевидно, причиной этого является образование билирубина типа III, моноконъюгированного, который ковалентно связан с альбумином. Он не фильтруется в клубочках и, следовательно, не появляется в моче. Это снижает практическую значимость проб, применяемых для определения содержания билирубина в моче.
Экскреция билирубина в канальцы происходит с помощью семейства АТФ-зависимых мультиспецифичных транспортных белков для органических анионов. Скорость транспорта билирубина из плазмы в жёлчь определяется этапом экскреции глюкуронида билирубина.
Жёлчные кислоты переносятся в жёлчь с помощью другого транспортного белка. Наличие разных механизмов транспорта билирубина и жёлчных кислот можно проиллюстрировать на примере синдрома Дубина-Джонсона, при котором нарушается экскреция конъюгированного билирубина, но сохраняется нормальная экскреция жёлчных кислот. Большая часть конъюгированного билирубина в жёлчи находится в смешанных мицеллах, содержащих холестерин, фосфолипиды и жёлчные кислоты. Значение аппарата Гольджи и микрофиламентов цитоскелета гепатоцитов для внутриклеточного транспорта конъюгированного билирубина пока не установлено.
Диглюкуронид билирубина, находящийся в жёлчи, водорастворим (полярная молекула), поэтому в тонкой кишке не всасывается. В толстой кишке конъюгированный билирубин подвергается гидролизу b-глюкуронидазами бактерий с образованием уробилиногенов. При бактериальном холангите часть диглюкуронида билирубина гидролизуется уже в жёлчных путях с последующей преципитацией билирубина. Этот процесс может иметь важное значение для образования билирубиновых жёлчных камней.
Уробилиноген, имея неполярную молекулу, хорошо всасывается в тонкой кишке и в минимальном количестве – в толстой. Небольшое количество уробилиногена, которое в норме всасывается, вновь экскретируется печенью и почками {энтерогепатическая циркуляция). При нарушении функции гепатоцитов печёночная реэкскреция уробилиногена нарушается и увеличивается почечная экскреция. Данный механизм объясняет уробилиногенурию при алкогольной болезни печени, при лихорадке, сердечной недостаточности, а также на ранних стадиях вирусного гепатита.
Источник