Оксигенация гемоглобина и миоглобина
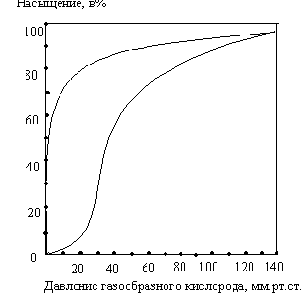
Особые свойства молекулы гемоглобина, которые делают его столь эффективным переносчиком кислорода в крови, легче всего уяснить из сравнения миоглобина и гемоглобина в отношении их сродства к кислороду. На рис. 8-16 показаны кривые насыщения кислородом для гемоглобина и миоглобина, характеризующие степень насыщения этих белков кислородом (т.е. отношение числа участков молекулы, связывающих кислород, к общему числу участков, способных к такому связыванию) в зависимости от парциального давления газообразного кислорода, находящегося в равновесии с раствором белка.
Рис. 8-15. Фоток рафия нормальных эритроцитов человека, полученная при помощи сканирующего электронного микроскопа.
Прежде всего из графика ясно, что миоглобин имеет очень высокое сродство к кислороду: при парциальном давлении кислорода, равном всего лишь 1-2 мм рт. ст., он уже на 50% насыщен кислородом. Кроме того, мы видим, что кривая насыщения миоглобина кислородом имеет вид простой гиперболы, как и следует ожидать из закона действующих масс применительно к равновесной реакции:
При парциальном давлении кислорода, равном 20 мм рт. ст., миоглобин оказывается насыщенным кислородом более чем на 95%. В отличие от миоглобина гемоглобин характеризуется значительно более низким сродством к кислороду; кроме того, кривая насыщения гемоглобина кислородом имеет сигмоидную, т.е. S-образную, форму (рис. 8-16). Это означает, что при связывании первой молекулы кислорода (нижняя часть S-образной кривой, соответствующая парциальным давлениям кислорода ниже 10 мм рт. ст.), гемоглобин имеет очень низкое сродство к кислороду, тогда как при связывании следующих молекул кислорода его сродство к ним становится намного выше, о чем свидетельствует крутая часть -образной кривой.
Рис. 8-16. Кривые насыщения кислородом для миог лобина и гемоглобина. Миоглобин обладает намного более высоким сродством к кислороду, чем гемоглобин. 50%-ное насыщение миоглобина кислородом достигается уже тогда, когда парциальное давление О, составляет всего 1 -2 мм рт. ст., тогда как для гемоглобина такое насыщение кислородом наступает лишь при парциальном давлении кислорода около 26 мм рт. ст. Обратите внимание, что в артериальной крови, вытекающей из легких (при парциальном давлении кислорода около 100 мм рт. ст.) оба белка – и миоглобин и гемоглобин – насыщены кислородом более чем на 95 %„ тогда как в покоящейся мыщце, где парциальное давление кислорода равно 40 мм рт. ст., гемоглобин насыщен кислородом лишь на 75%, а в работающей мышце при парциальном давлении кислорода всего около 10 мм рт. ст. только на 10%. Таким образом, гемоглобин очень эффективно отдает свой кислород в мышцах и других периферических тканях. Что же касается миоглобина, то при парциальном давлении кислорода, равном всего 10 мм рт. ст„ он все еще остается насыщенным кислородом почти на 90% и поэтому даже при столь низких парциальных давлениях кислорода отдает очень малую часть связанного с ним кислорода. Таким образом, сигмоидная кривая насыщения гемоглобина кислородом является результатом молекулярной адаптации гемоглобина к выполнению им транспортной функции в составе эритроцитов.
Фактически после связывания первой молекулы кислорода сродство повышается почти в 500 раз. Таким образом, четыре гемсодержащие полипептидные субъединицы гемоглобина различаются по степени их сродства к кислороду и зависят друг от друга в процессе его связывания.
Как только первая гемсодержащая полипептидная субъединица свяжет молекулу кислорода, она передает информацию об этом остальным субъединицам, у которых сразу же резко повышается сродство к кислороду. Такой обмен информацией между четырьмя гемсодержащими полипептидными субъединицами гемоглобина обусловлен кооперативным взаимодействием между субъединицами. Поскольку связывание первой молекулы кислорода одной из субъединиц гемоглобина увеличивает вероятность связывания следующих молекул кислорода остальными субъединицами, мы говорим, что гемоглобин имеет положительную кооперативностъ. Для положительной кооперативности характерны сигмоидные кривые связывания, подобные кривой насыщения гемоглобина кислородом. При связывании кислорода миоглобином, содержащим одну гемогруппу, молекула белка может присоединить только одну молекулу кислорода; в этом случае кооперативного связывания не наблюдается и кривая насыщения имеет вид простой гиперболы. Теперь мы понимаем. почему миоглобин и гемоглобин столь сильно различаются между собой по кислород-связывающей способности.
Мы будем использовать термин лиганд для обозначения специфической молекулы, связывающейся с белком; это может быть, например, молекула кислорода, если речь идет о гемоглобине (слово «лиганд» происходит от латинского слова, которое переводится как «связывать», «присоединять» и буквально означает «то, что присоединяется»). Многие другие олигомерные белки тоже имеют по нескольку лиганд-связывающих центров и, подобно гемоглобину, проявляют положительную кооперативность. Однако есть олигомерные белки, проявляющие отрицательную кооперативность: в этом случае связывание одной молекулы лиганда уменьшает вероятность связывания других молекул лиганда.
МИОГЛОБИН – сложный глобулярный белок, третьего уровня структурной организации, молекула которого состоит из 1 полипептидной цепи и содержит 153 аминокислоты. В миоглобине содержится железопорфириновая группа (гем), и он способен обратимо присоединять кислород.
Миоглобин содержится в клетках скелетных мышц. Пептидная цепь миоглобина напоминает длинную колбасу, причудливо скрученную. Было показано, что остов молекулы миоглобина состоит из 8 относительно прямолинейных отрезков, разделенных между собой местами сгибов. Каждый отрезок закручен в виде a-спирали. Все спирали являются правыми. 70 % аминокислотных остатков входят в состав спирализованных участков.
Свойства миоглобина:
1. молекула миоглобина компактна (внутри нее может уместиться 4Н2О);
2. все полярные R- группы аминокислотных остатков расположены на внешней поверхности молекулы и находятся в гидратированном состоянии, т.е. связаны с Н2О;
3. неполярные, или гидрофобные R- группы располагаются в глубине молекулы и защищены от соприкосновения с Н2О;
4. остатки пролина встречаются только в метах сгибов пептидной цепи (пролин нарушает a-спираль). В местах сгибов находятся и другие аминокислоты, которые неспособны легко образовывать a-спираль (изолейцин, серин), и аминокислоты, боковые цепи которых несут одинаковые заряды при рН7;
5. у миоглобинов, выделенных из разных млекопитающих, конформация пептидных цепей сходна (но они несколько отличаются по аминокислотному составу).
ГЕМ – комплекс порфирина и иона железа в степени окисления +2.
Ион железа встроен в кольцо порфирина таким образом, что четыре координационные связи из шести (в состоянии гибридизации sp3d2 связи у шестикоординационного железа направлены к вершинам октаэдра) затрачены на образование связей с атомами азота, еще одна связана с азотом имидазольного остатка ГИС полипептидной цепи (проксимальный Гистидин F8), а другая- также с имидазольным остатком другого ГИС (дистальный ГИС Е7). Молекула кислорода присоединяется между остатком дистального ГИС и железом. Изменения степени окисления железа при этом не происходит. Порфириновое кольцо (ГЕМ) не находится на плоскости молекулы белка, а частично погружено в него. Молекула кислорода присоединяется к гему, входя как бы через открывающуюся дверцу. Пока остается несным, дожидается молекула кислорода случайного открывания двери, или существует какой-то механизм, пускающий кислород к гему.
Миоглобин сосредоточен, главным образом, в мышцах и его главной функцией является хранение кислорода. Скорость насыщения миоглобина кислородом намного превышает таковую для гемоглобина. Миоглобин мало приспособлен для транспортировки кислорода из легких в ткани, поскольку скорость отдачи кислорода в тканях невелика (при давлении 1 мм рт. ст. примерно половина миоглобина все еще не отдает кислород).
Вопросы транспортировки кислорода решаются при участии белка четвертичной структуры – гемоглобина.
ЧЕТВЕРТИЧНАЯ СТРУКТУРА ГЕМОГЛОБИНА. При помощи рентгеноструктурного анализа Перутцем и его сотрудниками в Кембридже установлены третичная и четвертичная структуры гемоглобина. Гемоглобин содержится в эритроцитах и служит для переноса кислорода. Молекулярная масса гемоглобина 64500. Молекула состоит из 4 отдельных полипептидных цепей: 2 a-цепей (141 остаток аминокислот) и 2 b- цепей (146 остатков аминокислот в каждой), каждая из которых связана нековалентной связью с остатками гема. Каждая из 4 отдельных цепей гемоглобина свернута нерегулярным образом и состоит из ряда a- спиральных участков, разделенных местами сгибов.
a- и b- цепи гемоглобина примерно на 70 % состоят из a-спиральных участков. По своей третичной структуре a- и b-цепи очень сходны, они образованы из a- спиральных участков одинаковой длины, согнутых под одинаковыми углами и в одних и тех же направлениях. Третичная структура a- и b-цепей гемоглобина очень сходна с третичной структурой единственной цепи миоглобина. Сходная функция гемоглобина и миоглобина, обусловленная способностью обратимо связывать О2, объясняется сходством третичной структуры.
Согласно данным рентгеноструктурного анализа молекула гемоглобина по своей форме приближается к сфере диаметром ~ 5,5 нм. 4 полипептидные цепи уложены относительно друг друга приблизительно в виде тетраэдра, в результате чего возникает характерная четвертичная структура гемоглобина.
Это очень компактная структура. Большинство гидрофобных R- групп аминокислот находится внутри глобулы, а большинство гидрофильных R- групп – снаружи. В молекуле гемоглобина возникает небольшое число контактов между одинаковыми цепями (2 a- и 2 b- цепями) и множество контактов между a- и b- цепями. В образовании таких контактов принимают участие в основном гидрофобные R- группы аминокислотных остатков.
При присоединении к гемоглобину кислорода расстояние между 2 b- цепями гемоглобина уменьшается и изменяется четвертичная структура. Таким образом, гемоглобин и оксигемоглобин (насыщенный кислородом) различаются по своей четвертичной структуре.
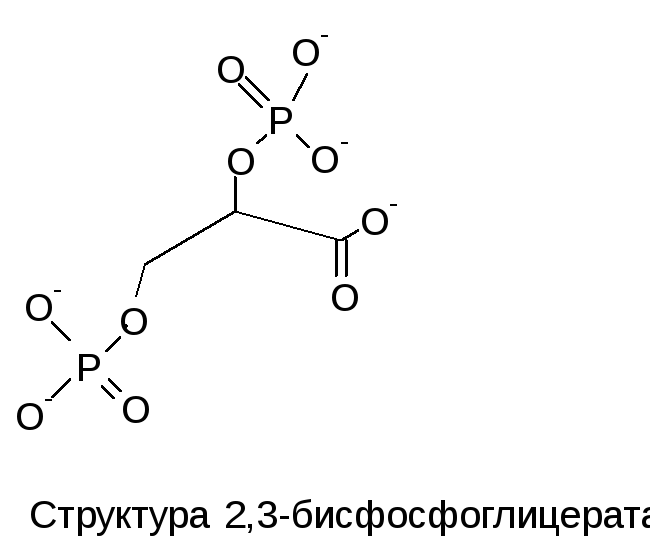
Четвертичная структура олигомерных белков также определяется первичной аминокислотной последовательностью входящих в их состав отдельных полипептидных цепей. Олигомерные белки (гемоглобин) обнаруживают способность к самосборке.
Главное отличие гемоглобина от миоглобина заключается в проявлении особого рода эффектов – кооперативных, влияющих на скорости присоединения- отсоединения молекул кислорода. Каждая молекула гемоглобина способна присоединять и переносить четыре молекулы кислорода, при этом кооперативность проявляется в том, что как присоединение, так и отсоединение каждой последующей молекулы кислорода облегчается в результате структурных изменений в конформации молекулы, которых у гемоглобина имеется две основных- оксигенированная и дезоксигенированная. Промежуточные состояния нестабильны. Предполагается следующий механизм кооперативного эффекта. Присоединение первой молекулы кислорода приводит к тому, что атом железа смещается от своего места примерно на 0,4-0,6 ангстрем, вызывая изменения конформации субъединицы. Изменившаяся конформация по аллостерическому эффекту облегчает присоединение кислорода к другой субъединице и т.д. Это позволяет максимально ускорить процесс присоединения кислорода в легких (рО2 = 100 мм рт. ст.). При переносе оксигенированного гемоглобина в капилляры тканей (рО2 = 5 мм рт. ст.) отсоединение молекул кислорода протекает также быстро, по кооперативному эффекту. Известны, впрочем, и химические регуляторы скорости и полноты присоединения кислорода. К ним, в частности, относится 2,3- дифосфоглицериновая кислота. Она облегчает присоединение кислорода у организмов, обитающих в высокогорных районах.
Оглавление темы “Вентиляция легких. Перфузия легких кровью.”: Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.Молекула гемоглобина может находиться в двух формах — напряженной и расслабленной. Расслабленная форма гемоглобина имеет свойство насыщаться кислородом в 70 раз быстрее, чем напряженная. Изменение фракций напряженной и расслабленной формы в общем количестве гемоглобина в крови обусловливает S-образный вид кривой диссоциации оксигемоглобина, а следовательно, так называемое сродство гемоглобина к кислороду. Если вероятность перехода от напряженной формы гемоглобина к расслабленной больше, то возрастает сродство гемоглобина к кислороду, и наоборот. Вероятность образования указанных фракций гемоглобина изменяется в большую или меньшую сторону под влиянием нескольких факторов. Основной фактор — это связывание кислорода с геминовой фуппой молекулы гемоглобина. При этом чем больше геминовых фупп гемоглобина связывают кислород в эритроцитах, тем более легким становится переход молекулы гемоглобина к расслабленной форме и тем выше их сродство к кислороду. Поэтому при низком Р02, что имеет место в метаболически активных тканях, сродство гемоглобина к кислороду ниже, а при высоком Р02 — выше. Как только гемоглобин захватывает кислород, повышается его сродство к кислороду и молекула гемоглобина становится насыщенной при связывании с четырьмя молекулами кислорода.
Когда эритроциты, содержащие гемоглобин, достигают тканей, то кислород из эритроцитов диффундирует в клетки. В мышцах он поступает в своеобразного депо кислорода — в молекулы миоглобина, из которого кислород используется в биологическом окислении мышц. Диффузия кислорода из гемоглобина эритроцитов в ткани обусловлена низким Р02 в тканях — 35 мм рт. ст. Внутри клеток тканей напряжение кислорода, необходимое для поддержания нормального метаболизма, составляет еще меньшую величину — не более 1 кПа. Поэтому кислород путем диффузии из капилляров достигает метаболически активных клеток. Некоторые ткани приспособлены к низкому содержанию Р02 в капиллярах крови, что компенсируется высокой плотностью капилляров на единицу объема тканей. Например, в скелетной и сердечной мышцах Р02 в капиллярах может снизиться чрезвычайно быстро во время сокращения. В мышечных клетках содержится белок миоглобин, который имеет более высокое сродство к кислороду, чем гемоглобин. Миоглобин интенсивно насыщается кислородом и способствует его диффузии из крови в скелетную и сердечную мышцы, где он обусловливает процессы биологического окисления. Эти ткани способны экстрагировать до 70 % кислорода из крови, проходящей через них, что обусловлено снижением сродства гемоглобина к кислороду под влиянием температуры тканей и рН. Эффект рН и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое рН понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и С02. Если рН плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном рН 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда рН снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 1/з больше, чем при постоянном рН.
Метаболически активные ткани повышают продукцию тепла. Повышение температуры тканей при физической работе изменяет соотношение фракций гемоглобина в эритроцитах и вызывает смещение кривой диссоциации оксигемоглобина вправо вдоль оси х. В результате большее количество кислорода будет освобождаться из гемоглобина эритроцитов и поступать в ткани. Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении Р02 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3-ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей. – Также рекомендуем “Углекислый газ. Транспорт углекислого газа.” |


Почему
миоглобин не способен транспортировать
кислород, но зато активно его запасает?
Для миоглобина изотерма адсорбции
кислорода имеет форму гиперболы. Давление
О2
в ткани, окружающей легочные капилляры,
составляет 100 мм рт.ст., поэтому миоглобин
в легких мог бы весьма эффективно
насыщаться кислородом. В венозной крови
Р О2
равно 40 мм рт. ст., а в активно работающей
мышце – 20 мм рт.ст. Но даже при парциальном
давлении 20 мм рт. ст. степень насыщения
миоглобина кислородом будет весьма
значительна, и поэтому миоглобин не
может служить транспортной молекулой
для доставки О2
от легких к периферическим тканям.
Однако при кислородном голодании,
которым сопровождается тяжелая физическая
работа, парциальное давление О2
в тканях может понизиться и до 5 мм
рт.ст.; при столь низком давлении миоглобин
легко отдает кислород, обеспечивая тем
самым окислительный синтез АТФ в
митохондриях мышечных клеток.
Кинетика
оксигенирования гемоглобина коренным
образом отличается от кинетики
оксигенирования миоглобина. Кривая
насыщения гемоглобина О2
имеет сигмоидальную форму. Т.о., способность
гемоглобина связывать О2
зависит от того, содержатся ли в данном
тетрамере другие молекулы О2.
Если да, то последующие молекулы О2
присоединяются легче. Для гемоглобина
характерна кинетика кооперативного
связывания, благодаря которой он
связывает максимальное количество О2
в легких и отдает максимальное количество
О2
при тех значениях Р О2
, которые имеют место в периферических
тканях.
Сродство
гемоглобинов к О2
характеризуется величиной Р50-
значением парциального давления О2,
при котором наблюдается полунасыщение
гемоглобина кислородом. Например, для
HbA
Р50
= 26 мм рт.ст., а для HbF
– 20 мм рт. ст. Благодаря этой разнице
гемоглобин F
отбирает кислород у HbA,
находящегося в плацентарной крови.
2)
У олигомерных белков имеется специальный
центр, отличный от активного, к которому
присоединяется аллостерический лиганд.
Этот центр получил название аллостерического.
Рассмотрим
аллостерическую регуляцию на примере
HbА.
В центре молекулы HbА
находится полость, которая является
местом присоединения 2,3-ДФГ. Это соединение
накапливается в эритроцитах при
недостатке О2.
Непосредственным предшественником
служит 1,3-дифосфоглицерат – промежуточный
продукт гликолиза. 2,3-ДФГ имеет сильный
отрицательный заряд и взаимодействует
с 5 положительно заряженными группами
аллостерического центра. В результате
взаимодействия образуется 5 дополнительных
ионных связей, что снижает сродство Hb
к кислороду и облегчает его отдачу в
ткани.
Кооперативные
изменения конформации олигомерных
белков составляют основу механизма
регуляции функциональной активности
не только Hb,
но и большого числа других белков, в том
числе аллостерических
ферментов.
5. Изофункциональные белки
Белок,
выполняющий определенную функцию в
клетке, может быть представлен несколькими
формами – изофункциональными белками
или изобелками.
Такие белки хоть и выполняют одинаковую
функцию, но отличаются константой
связывания, что приводит к некоторым
различиям в функциональном отношении.
Например, в эритроцитах человека
обнаружено несколько форм гемоглобина:
HbA
(96%), HbF
(2%), HbA2
(2%).
Все гемоглобины представляют собой
тетрамеры, построенные из протомеров
,
,
,
(HbA
-22,
HbF
– 22,
HbA2
– 22).
Все протомеры сходны между собой по
первичной, вторичной и третичной
структурам. Все формы гемоглобинов
предназначены для переноса кислорода
в клетки тканей, но HbF,
например, имеет большее сродство к
кислороду, чем HbA.
HbF
характерен для эмбриональной стадии
развития человека. Он способен отнимать
кислород у HbA,
что обеспечивает нормальное снабжение
кислородом плода.
Изобелки
– это результат наличия более чем одного
структурного гена в генофонде вида.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #