При разрушении гемоглобина образуются пигменты

Гемоглобин (высокомолекулярный хромопротеид) – железосодержащий дыхательный пигмент, составляющий основу эритроцитов и выполняющий роль носителя кислорода. В результате физиологического распада (гемолиза) эритроцитов и гемоглобина образуются пигменты: 1) ферритин (- железопротеид, содержащий до 23% железа; в зависимости от происхождения различают анаболический (образуется из железа, всасывающегося в кишечнике) и катаболический (образуется из железа гемолизированных эритроцитов) ферритин; обладает антигенными свойствами; образует берлинскую лазурь (железосинеродистое железо); большое количество ферритина содержится в печени (депо ферритина), селезенке, костном мозге и лимфатических узлах, где обмен его связан с синтезом гемосидерина, гемоглобина и цитохромов;
в условиях патологии количество ферритина может увеличиваться как в крови, так и в тканях; повышение содержания ферритина в тканях набл-ся при гемосидерозе, т.к. полимеризация ферритина ведет к обр-ю гемосидерина). 2) гемосидерин (- образуется при расщеплении гема и является полимером ферритина; клетки, в которых образуется гемосидерин, называются сидеробластами; гемосидерин обнаруживается в ретикулярных и эндотелиальных клетках селезенки, печени, костного мозга, лимфатических узлов; в межклеточном веществе он подвергается фагоцитозу сидерофагами; в условиях патологии набл-ся избыточное образование гемосидерина – гемосидероз – м.б. как общим (наб-ся при внутрисосудистом разрушении эритроцитов – интраваскулярный гемолиз), так и местным (развивается при внесосудистом разрушении эритроцитов – экстраваскулярный гемолиз)). 3) билирубин(- желчный пигмент; он не содержит ни железа, ни белка; формируется в гистиоцитарно-макрофагальной системе; транспортируется кровью в печень в несвязанной форме (непрямой, или несвязанный билирубин – растворим в липидах) в комплексе с альбумином (прямой, или связанный билирубин). В печени билирубин ферментативно связывается с глюкуроновой кислотой, формируя водорастворимый прямой (связанный) билирубин, который экскретируется клетками печени в желчь, а затем попадает в кишечник (холебилирубин). В кишечнике благодаря бактериальной активности он преобразовывается в уробилиноген; в норме билирубин встречается в растворенном состоянии в желчи и в небольшом кол-ве в плазме крови; нарушение обмена билирубина связано с расстройством его образования и выделения —- ведет к повышению содержания в плазме крови с накоплением его в тканях и желтушным окрашиванием кожи, склер, слизистых, серозных оболочек и внутренних органов — желтуха). В патологических условиях, помимо увеличения образующихся в норме гемоглобиногенных пигментов, может появляться ряд новых пигментов – 1) гематоидин (- не содержит железа; возникает при распаде эритроцитов и гемоглобина внутриклеточно, спустя 5-10 дней после гемосидерина, но в отличие от гемосидерина в клетках не остается и при их гибели оказывается свободно лежащим среди некротических масс; скопления гематоидина находят в более или менее старых гематомах, рубцующихся инфарктах, причем вдали от живых тканей – в центральных участках кровоизлияний, а не по периферии; его образование связано с распадом клеток, т.е. с угасанием и прекращением жизнедеятельности в очаге кровоизлияния при недостаточном доступе кислорода). 2) гематины (- окисленная форма гема; образуются при гидролизе оксигемоглобина; содержат железо в связанном состоянии; к гематинам относят – гемомеланин (малярийный пигмент); солянокислый гематин (гемин); формалиновый пигмент; гистохимические свойства этих пигментов идентичны). 3) порфирин (-флюоресцирующий пигмент, близкий по структуре билирубину, содержит железо; в норме в небольшом количестве содержится в крови и в моче, играет роль антагониста меланина и повышает чувствительность кожи к свету; при нарушении его обмена –увеличивается содержание порфирина в крови (порфиринемия) и в моче (порфиринурия); моча, содержащая большое количиство порфирина, становится красной; пигмент обнаруживается также и в кале. Это состояние носит название порфирия. Причины порфирии: интоксикации (отравления свинцом, сульфоналом, барбитуратами); авитаминоз РР (при пеллагре); врожденные дефекты нарушения обмена; при порфирии развиваются более или менее тяжелые симптомы повышенной чувствительности кожи к ультрафиолетовым лучам; на открытых частях тела – на лице, на руках, шее возникают эритемы, пузыри, язвы на фоне глубокой атрофии кожи с ее депигментацией; кости и зубы становятся коричневыми).
Источник
Разрушение гемоглобина. Разновидности анемий
При разрыве эритроцитов их гемоглобин почти сразу же фагоцитируется макрофагами во многих частях тела, но особенно клетками Купфера печени и макрофагами селезенки и костного мозга. В течение нескольких следующих часов или дней макрофаги освобождают железо из гемоглобина, и оно возвращается в кровь и переносится трансферрином либо в костный мозг для формирования новых красных клеток крови, либо в печень и другие ткани для хранения в форме ферритина.
Порфириновая часть молекулы гемоглобина превращается макрофагами через ряд стадий в желчный пигмент билирубин, который выделяется в кровь и позднее удаляется из организма путем секреции печенью в желчь.
Анемия означает недостаток гемоглобина в крови, причиной может быть либо слишком малое число красных клеток крови, либо слишком малое количество гемоглобина в этих клетках. Далее представлены некоторые типы анемий и их физиологические причины.
Анемия, связанная с потерей крови. После острой кровопотери организм возмещает жидкую часть плазмы в течение 1-3 сут, но при этом концентрация красных клеток крови остается низкой. Концентрация эритроцитов обычно восстанавливается до нормы в течение 3-6 нед, если не происходит повторного кровотечения.
Часто при хронической кровопотере у человека железо из кишечника не может всасываться достаточно быстро, чтобы обеспечить адекватное возмещение теряемого с кровью гемоглобина. Формируемые в этом случае красные клетки крови гораздо мельче нормальных эритроцитов и содержат слишком мало гемоглобина, что характерно для микроцитарной гипохромной анемии; такие эритроциты показаны на рисунке.
Апластическая анемия. Аплазия костного мозга означает потерю функционирующего костного мозга. Например, у человека, подвергшегося облучению гамма-лучами при взрыве атомной бомбы, может произойти полное разрушение костного мозга с последующим развитием в течение нескольких недель летальной анемии. Тот же эффект могут вызвать избыточная рентгенотерапия, некоторые промышленные химикаты и даже лекарства, к которым у человека может быть повышенная чувствительность.
Мегалобластная анемия. На основании изложенного ранее обсуждения роли витамина B12, фолиевой кислоты и внутреннего фактора, секретируемого слизистой желудка, легко понять, что недостаток любого из этих веществ может привести к замедлению репродукции эритроцитов в костном мозге. В результате формируются слишком крупные красные клетки крови разнообразной формы, которые называют мегалобластами.
Следовательно, атрофия слизистой желудка, например при пернициозной анемии, или потеря всего желудка после хирургической тотальной гастрэктомии могут привести к мегалобластной анемии. Мегалобластная анемия часто развивается также у больных с кишечной спру, при которой плохо всасываются фолиевая кислота, витамин B12 и другие соединения витаминов группы В. Поскольку при этих состояниях эритробласты не могут пролиферировать достаточно быстро, чтобы формировать нормальное количество красных клеток крови, те эритроциты, которые формируются, по большей части увеличены в размерах, имеют неправильную форму и ломкие мембраны. Эти клетки легко рвутся, оставляя человека без необходимого количества красных клеток крови.
Гемолитическая анемия. Различные аномалии красных клеток крови, многие из которых — наследственные, делают клетки столь хрупкими, что они легко разрываются, проходя через капилляры, особенно в селезенке. Даже если количество формируемых красных клеток крови в норме или значительно ее превышает, как при некоторых гемолитических болезнях, срок жизни хрупкого эритроцита так короток, что клетки разрушаются быстрее, чем могут формироваться нормальные эритроциты; результатом этого является тяжелая анемия. Далее указаны некоторые из таких типов анемий.
При наследственном сфероцитозе красные клетки крови очень мелкие и сферические, а не двояковогнутые диски. Эти клетки не могут выдерживать сдавливания, поскольку не имеют нормальной свободной, мешкообразной клеточной мембраны, характерной для двояковогнутых дисков. При прохождении через пульпу селезенки и некоторые другие сосудистые ложа они легко ломаются даже при небольшом сдавливании.
При серповидно-клеточной анемии, которой болеют 0,3-1,0% коренных жителей Западной Африки, красные клетки крови содержат аномальный гемоглобин — гемоглобин S с поврежденными цепочками в его молекуле. Под действием низких концентраций кислорода такой гемоглобин осаждается в виде длинных кристаллов внутри эритроцита. Эти кристаллы удлиняют клетку и придают ей вид серпа, а не двояковогнутого диска.
Кроме того, осажденный гемоглобин повреждает клеточную мембрану, в результате клетка становится очень хрупкой, что сопровождается тяжелой анемией. Такие больные часто переживают порочный круг событий, называемый кризисом серповидно-клеточной болезни, при котором низкое напряжение кислорода в тканях вызывает образование серповидных форм эритроцитов, что ведет к разрушению красных клеток крови, а значит — к дальнейшему снижению напряжения кислорода, усилению образования серповидных форм и разрушению красных клеток крови. Сразу после начала процесс быстро прогрессирует, приводя в течение нескольких часов к резкому снижению числа красных клеток крови и часто — к смерти.
При гемолитической болезни новорожденных (эритробластозе) антитела от резус-отрицательной (Rh-) матери атакуют резус-положительные (Rh+) эритроциты плода. В результате резус-положительные клетки становятся ломкими, что ведет к их быстрому разрушению, способствуя развитию у новорожденного тяжелой анемии. Чрезвычайно быстрое формирование новых эритроцитов для возмещения разрушенных при гемолитической болезни новорожденных ведет к выделению в кровь из костного мозга большого количества молодых бластных форм красных клеток крови.
– Также рекомендуем “Влияние анемии на кровообращение. Полицитемия – эритремия”
Оглавление темы “Эритропоэз. Белые клетки крови”:
1. Влияние эритропоэтина на эритрогенез. Витамин В12 и фолиевая кислота в эритропоэзе
2. Пернициозная анемия. Образование гемоглобина
3. Связывание гемоглобина с кислородом. Обмен железа
4. Всасывание железа в кишечнике. Длительность жизни эритроцитов
5. Разрушение гемоглобина. Разновидности анемий
6. Влияние анемии на кровообращение. Полицитемия – эритремия
7. Влияние полицитемии на кровообращение. Лейкоциты – белые клетки крови
8. Типы белых клеток крови. Происхождение белых клеток крови
9. Длительность жизни белых клеток крови. Нейтрофилы и макрофаги
10. Фагоцитоз. Механизмы и значение фагоцитоза
Источник
За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов.
Эритроциты в норме живут 90-120 дней, после чего лизируются в клетках ретикулоэндотелиальной системы – макрофагах селезенки (главным образом), купферовских клетках печени и макрофагах костного мозга. При разрушении эритроцитов в кровеносном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и также переносится в клетки РЭС селезенки, печени и костного мозга.
Синтез билирубина
В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина. Высвобождаемое железо может либо запасаться в клетке в комплексе с ферритином, либо выделяться наружу и связываться с трансферрином.
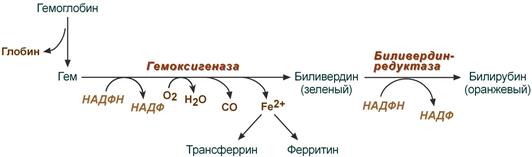
Реакции распада гемоглобина и образования билирубина
Билирубин – токсичное, жирорастворимое вещество, способное разобщать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
Строение билирубина
Выведение билирубина
Из клеток ретикуло-эндотелиальной системы билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.
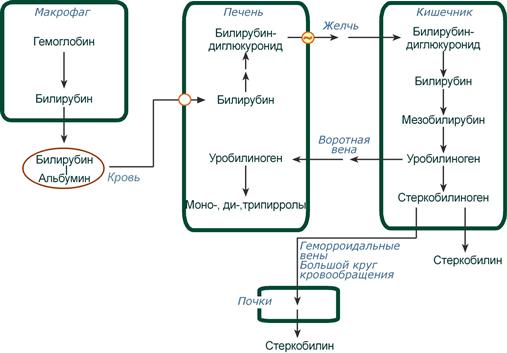
Этапы метаболизма билирубина в организме
Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (транспортный белок органических анионов) или по механизму флип-флоп. Далее при участии цитозольного связывающего белка лигандина (Y-протеин) билирубин транспортируется в ЭПР, где протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию конъюгации могут вступать сульфаты, фосфаты, глюкозиды.
Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин. Одновременно, даже в норме (особенно у взрослых), некоторое количество билирубин-глюкуронидов может попадать из желчи в кровь по межклеточным щелям.
Таким образом, в плазме крови обычно присутствуют две формы билирубина: свободный (непрямой), попадающий сюда из клеток РЭС (80% и более всего количества), и связанный (прямой), попадающий из желчных протоков (в норме не более 20%).
Термины “связанный“, “конъюгированный“, “свободный“, “несвязанный” отражают взаимодействие билирубина и глюкуроновой кислоты (но не билирубина и альбумина!).
Термины “прямой” и “непрямой” введены, исходя из возможности химической реакции билирубина с диазореактивом Эрлиха. Связанный билирубин реагирует с реактивом напрямую, без добавления дополнительных реагентов, т.к. является водорастворимым. Несвязанный (жирорастворимый) билирубин требует добавочных реактивов, реагирует не прямо.
Превращение в кишечнике
В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть уробилиногена всасывается и с кровью портальной вены попадает в печень, где либо распадается до моно-, ди- и трипирролов, либо окисляется до билирубина и снова экскретируется. При этом при здоровой печени в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена. Далее
- малая часть стеркобилиногена может всасываться и катаболизировать в печени, подобно уробилиногену,
- незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и в мочу. После окисления на воздухе из стеркобилиногена образуется стеркобилин мочи,
- однако основное количество стеркобилиногена достигает нижних отделов толстого кишечника и выделяется. В прямой кишке и на воздухе стеркобилиноген окисляется в стеркобилин, окрашивая кал,
- аналогично уробилиноген, появляющийся в моче при патологии печени, окисляется в уробилин.
Очень часто стеркобилиноген, содержащийся в нормальной моче, называют уробилиногеном. И в клинической практике обычно не проводят различий между стеркобилиногеном и уробилиногеном мочи, их рассматривают как один пигмент – урохромы (уробилиноиды), что может создавать некоторую путаницу при оценке результатов анализа.
Источник
Схема 8. Циклы железа в организме
ГЕМОГЛОБИНОГЕННЫЕ ПИГМЕНТЫ
ЭНДОГЕННЫЕ ПИГМЕНТАЦИИ
• Эндогенные пигментации — разновидность смешанных дистрофий. В основе их лежат нарушения эндогенных пигментов.
Эндогенные пигменты— окрашенные вещества различной химической природы, которые синтезируются в самом организме, придавая органам и тканям различную окраску. По своей структуре они являются хромопротеидами (от греч. chroma — цвет, окраска + протеиды), т.е. окрашенными белками. Хромопротеиды широко распространены в живой природе и выполняют разнообразнейшие биологические функции: перенос и депонирование кислорода для осуществления окислительно-восстановительных процессов в клетках, в том числе и дыхания (гемоглобин, цитохромы, миоглобин, липофусцин), рецепция света и защита от действия ультрафиолетового излучения (меланин), синтез биологически активных веществ (пигмент гранул энтерохромаффинных клеток), секретов (желчь), доставка и регуляция обмена микроэлементов (церулоплазмин, ферритин, гемосидерин), витаминов (липохромы) и др.
Классификация.Эндогенные пигменты разделяют, согласно их формальному генезу, на 3 группы:
▲ гемоглобиногенные, представляющие собой различные производные гемоглобина;
▲ протеиногенные, или тирозиногенные, связанные с обменом тирозина;
▲ липидогенные, или липопигменты, образующиеся при обмене жиров.
Продукты нарушенного обмена эндогенных пигментов обычно откладываются как в паренхиме органов, так и вне ее, в строме. При нарушении обмена пигментов учитывают следующие особенности:
количество пигмента. Оно может быть увеличено или, наоборот, уменьшено вплоть до полного исчезновения;
распространенность процесса (общий или местный характер процесса);
характер наследования. Этиологические факторы, вызывающие нарушение обмена хромопротеидов, являются генетически обусловленными или же приобретаются в течение жизни; в связи с этим различают наследственные и приобретенные нарушения обмена пигментов.
Пигментный обмен может нарушаться при многих болезнях и патологических состояниях, т.е. возникает вторично; однако иногда нарушения обмена хромопротеидов возникают первично и являются морфологическим субстратом самостоятельных заболеваний. В большинстве случаев патологические пигментации возникают в связи с избыточным накоплением пигментов, которые встречаются и в норме, но иногда накапливается пигмент, который возникает только в условиях патологии.
Гемоглобиногенные пигменты получили свое наименование вследствие того, что их образование связано с метаболизмом гемоглобина. При этом часть пигментов образуется в физиологических условиях. Это гемосидерин, ферритин и билирубин. Часть пигментов — гематоидин, гематины и порфирин, образуются только в условиях патологии. Некоторые из этих пигментов (ферритин, гемосидерин) синтезируются, помимо гемоглобина, из железа, всасывающегося в кишечнике (схема 8). Поэтому определение “гемоглобиногенные пигменты” является для них весьма условным.
Гемоглобин— хромопротеид, который в качестве простетической группы содержит железопорфириновый комплекс гем. Белковая часть молекулы гемоглобина состоит из двух пар полипептидных (а и Ь) цепей, содержащих по 140 аминокислот. Своим огромным значением гемоглобин обязан содержащемуся в нем железу, с которым филогенетически связана функция дыхания. Обмен гемоглобина тесно связан с эритроцитами, в которых он содержится, с их состоянием, старением, разрушением. Физиологический гемолиз происходит в основном в костном мозге, реже — в селезенке и печени, в клетках макрогистиоцитарной системы этих органов образуются ферритин, гемосидерин и билирубин.
Ферритин— железопротеид, содержащий белок апоферритин и трехвалентный атом железа в составе фосфатного гидроксида. Ферритин неоднороден, известно до 20 изоферритинов. Это разнообразие обусловлено различием вариантов входящего в его состав апоферритина (Н-, L- и HL-субъединицы), различием способов происхождения пигмента (“анаболический” — из железа, всасывающегося в кишечнике, “катаболический” — из железа гемолизированных эритроцитов), разной локализацией (в сыворотке крови — HL-ферритин, в печени и селезенке — L-ферритин). Наконец, важное значение имеет кислород: ферритин синтезируется из двухвалентного железа в присутствии кислорода и содержит много SS-групп. При гипоксии образуется SH-ферритин, обладающий вазопаралитическим действием. Значение ферритина трудно переоценить. Он является главным участником метаболизма железа. Известно, что свободные атомы железа токсичны для организма. Именно в форме ферритина депонируется железо (до 30 %, хотя расходуется только 0,1 %). Ферритин содержится практически во всех органах и тканях и является акцептором железа в клетках, которые в нем нуждаются (эритробласты). Он также осуществляет перенос железа в кишечнике и плаценте, т.е. является медиатором при соединении железа с трансферрином и в переносе его от матери к плоду.
Ферритин выявляют в тканях с помощью сульфата кадмия по методу Клочкова, а также иммуногистохимически с использованием специфических антисывороток. На практике чаще всего используется гистохимический метод — реакция образования берлинской лазури (железистосинеродистое железо) или реакции Перльса — реакция на выявление солей оксида железа (III) с помощью железосинеродистого калия и хлороводородной (соляной) кислоты.
Гемосидерин— это продукт полимеризации ферритина. По химической структуре он является коллоидным гидроксидом железа, соединенным с мукопротеидами клетки. В норме гемосидерин образуется в ретикулярных и эндотелиальных клетках селезенки, лимфатических узлов, печени и костного мозга. При окраске гематоксилином и эозином гемосидерин выявляется в виде зерен бурого цвета в цитоплазме этих клеток, а при реакции Перльса — в виде гранул зеленовато-синего цвета (берлинская лазурь). Гемосидерин — внутриклеточный пигмент. Синтез его происходит в клетках, которые называют сидеробластами, в специализированных органеллах — сидеросомах. Иногда в сидеробластах накапливается такое большое количество гемосидерина, что клетки разрушаются и гемосидерин оказывается свободно лежащим в строме органов. В этих случаях он обычно захватывается макрофагами, которые принято называть сидерофагами. В цитоплазме этих клеток сидеросомы не выявляются.
Билирубин— конечный продукт гемолиза. Билирубин образуется, когда от гемоглобина отщепляется гем, а затем от гема отщепляется железо и разворачивается тетрапиррольное кольцо. Этот процесс начинается в клетках ретикуломакрофагальной системы костного мозга, селезенки, лимфатических узлов и печени. Затем продукт, соединяясь с альбумином, с током крови поступает в печень. В печени синтез пигмента завершается — гепатоциты, обладая специфическими рецепторами, захватывают его и с помощью ферментов специфической глюкуронилтрансферазной системы осуществляют его конъюгацию. Конъюгаты билирубина поступают в желчные капилляры. Таким образом, билирубин становится основным пигментом желчи.
Обычно билирубин находится в виде кристаллов красновато-желтого цвета. Он легко окисляется, образуя при этом продукты различного цвета. Именно это происходит при выявлении его по методу Гмелина — при окислении его азотной кислотой образуются продукты сначала зеленого, а затем синего или пурпурного цвета.
Гематоидин— пигмент, не содержащий железа. По химической структуре близок к билирубину и также дает положительную реакцию Гмелина. Гематоидин формирует ярко-оранжевые кристаллы в виде ромбических пластинок, иголок или зерен. Образуется при распаде эритроцитов и гемоглобина, как и гемосидерин, внутриклеточно, но в клетках не остается и при их гибели оказывается свободно лежащим среди некротических масс.
Гематиныобразуются при гидролизе оксигемоглобина и представляют собой окисленную форму гема, содержащую трехвалентный атом железа в связанном состоянии. Имеют вид темно-коричневых кристаллов или зерен. К гематинам относят малярийный пигмент (гемомеланин), солянокислый гематин и формалиновый пигмент.
Малярийный пигмент (гемомеланин) образуется из гема в теле малярийного плазмодия, который, как известно, паразитирует в эритроцитах. Пигмент построен из буровато-черных аморфных гранул и синтезируется обычно в ретикулярных и эндотелиальных клетках печени, костного мозга, селезенки и лимфатических узлов.
Солянокислый гематин (гемин) образуется исключительно в желудке при взаимодействии гемоглобина, ферментов желудочного сока и соляной кислоты. Пигмент откладывается в виде ромбовидных или игловидных кристаллов.
Формалиновый пигмент образуется в тканях при фиксации их кислым формалином (рН<5,6), имеет вид бурых зерен или кристаллов, расположенных, как правило, в просвете венозных сосудов.
Порфирины — предшественники гема, которые имеют строение замкнутых тетрапиррольных колец, лишенных железа. Пигменты повышают чувствительность кожи к ультрафиолетовому облучению, являются антагонистами меланина. Обычно метаболизм порфиринов в организме человека заканчивается на стадии уропорфириногена III, который затем принимает участие в реакциях синтеза гема. При отсутствии фермента уропорфириноген Ш-косинтетазы появляются предшественники уропорфириногена III — уропорфириноген I, порфобилин, порфобилиногены. В норме они в минимальных количествах определяются в тканях, крови и моче: они дают оранжевую флюоресценцию в ультрафиолетовом свете.
Источник