Роль гемоглобина в транспорте газов крови

Гемоглобин. Роль гемоглобина в транспорте кислородаОбычно из легких в ткани переносятся эритроцитами в химической связи с гемоглобином около 97% кислорода. Оставшиеся 3% кислорода транспортируются в физической растворенной форме плазмой крови. Таким образом, в нормальных условиях почти весь кислород переносится в ткани, будучи связанным с гемоглобином. а) Обратимая связь кислорода с гемоглобином. Химический состав гемоглобина представлен в отдельных статьях на сайте, где говорилось, что молекула кислорода легко и обратимо связывается с гемом гемоглобина. При высоком PO2, как это бывает в легочных капиллярах, кислород связывается с гемоглобином, а при низком PO2, как в капиллярах тканей, кислород освобождается от связи с гемоглобином. Такой механизм обеспечивает почти весь транспорт кислорода из легких в ткани.
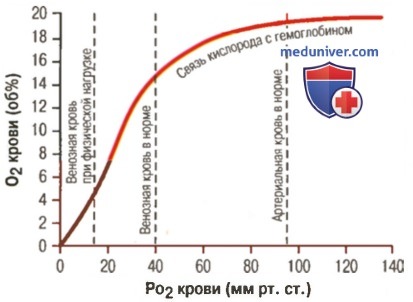
1. Кривая диссоциации оксигемоглобина. На рисунке выше приведена кривая диссоциации оксигемоглобина, демонстрирующая прогрессивный прирост процентной доли оксигемоглобина (процента насыщения гемоглобина кислородом) при увеличении PO2 в крови. В крови, покидающей легкие и входящей в системные артерии, напряжение О2 обычно составляет примерно 95 мм рт. ст., и на кривой диссоциации видно, что насыщение системной артериальной крови кислородом составляет 97%. В нормальной возвращающейся из периферических тканей венозной крови напряжение О2 составляет около 40 мм рт. ст. и 75% — насыщение гемоглобина кислородом. 2. Максимальное количество кислорода, которое может находиться в связи с гемоглобином крови. В 100 мл крови здорового человека содержится около 15 г гемоглобина, и каждый грамм гемоглобина может связать максимально 1,34 мл кислорода (химически чистый гемоглобин может связать 1,39 мл кислорода, но примеси типа метгемоглобина снижают это количество). Итак, 15×1,34 = 20,1, значит, в среднем содержащееся в 100 мл количество гемоглобина при 100% насыщении может связать около 20 мл кислорода. Обычно это обозначают как 20 об% (объемных процентов). Кривая диссоциации оксигемоглобина может строиться не от процентного насыщения гемоглобина кислородом, а от количества содержания объемных процентов кислорода. 3. Количество кислорода, высвобождаемого гемоглобином во время прохождения артериальной крови через ткани. В обычных условиях в системной артериальной крови, насыщенной кислородом на 97%, общее количество связанного с гемоглобином кислорода составляет около 19,4 мл на 100 мл крови (для облегчения понимания просим вас изучить рисунок ниже). При прохождении через капилляры ткани этот объем снижается до 14,4 мл (PO2 — 40 мм рт. ст., насыщение гемоглобина — 75%). Итак, в нормальных условиях каждые 100 мл крови доставляют от легких к тканям около 5 мл кислорода.
4. Транспорт кислорода при тяжелой физической нагрузке. При тяжелой физической работе мышечные клетки потребляют кислород с повышенной скоростью, что может привести к падению PO2 в интерстициальной жидкости мышцы от нормального уровня 40 мм рт. ст. до 15 мм рт. ст. При таком низком парциальном давлении в каждых 100 мл крови остается только 4,4 мл связанного с гемоглобином кислорода. В этом случае каждые 100 мл протекающей крови отдают тканям 19,4 – 4,4, или 15 мл кислорода, т.е. каждый объем крови отдает тканям в 3 раза больше кислорода, чем в покое. Вспомните, что у хорошо тренированных бегунов-марафонцев сердечный выброс может увеличиться в 6-7 раз, чем при покое. Если умножить это увеличение сердечного выброса (6-7 раз) на увеличение высвобождения кислорода в тканях каждым объемом крови (3 раза), получается, что к тканям было доставлено в 20 раз больше кислорода, чем в покое. Далее в этой главе Вы узнаете о существовании других факторов, улучшающих доставку кислорода в ткани во время физической нагрузки, поэтому даже при очень напряженной физической работе наблюдается только очень небольшое снижение PO2 в мышечной ткани. – Также рекомендуем “Коэффициент использования кислорода. Сохранение постоянства кислорода в тканях” Оглавление темы “Кислород и его доставка в организме”: |
Оглавление темы “Вентиляция легких. Перфузия легких кровью.”:
1. Вентиляция легких. Вентиляция кровью легких. Физиологическое мертвое пространство. Альвеолярная вентиляция.
2. Перфузия легких кровью. Влияние гравитации на вентиляцию легких. Влияние гравитации на перфузию легких кровью.
3. Коэффициент вентиляционно-перфузионных отношений в легких. Газообмен в легких.
4. Состав альвеолярного воздуха. Газовый состав альвеолярного воздуха.
5. Напряжение газов в крови капилляров легких. Скорость диффузии кислорода и углекислого газа в легких. Уравнение Фика.
6. Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
7. Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.
8. Углекислый газ. Транспорт углекислого газа.
9. Роль эритроцитов в транспорте углекислого газа. Эффект Холдена..
10. Регуляция дыхания. Регуляция вентиляции легких.
Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл • л-1 • кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
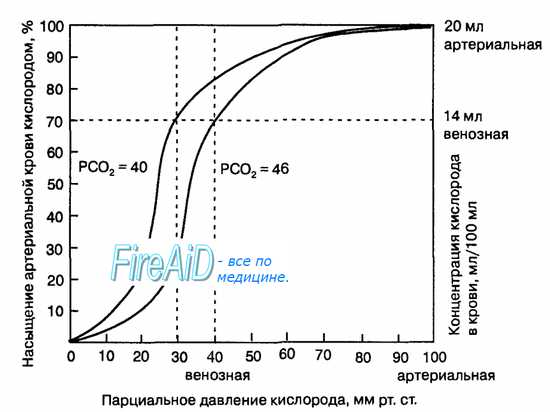
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
– Также рекомендуем “Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.”
Одной из основных функций, которые выполняет кровь в организме животного и человека, является дыхательная функция. Кровь, осуществляя транспорт кислорода воздуха от капилляров легочных альвеол к тканевым капиллярам, обеспечивает таким образом бесперебойную доставку кислорода, необходимого для нормального течения жизненно важных процессов в организме.
Переносимый кислород находится в крови в двух состояниях. Небольшая часть кислорода находится в растворенном состоянии в плазме. Коэффициент растворимости кислорода в крови невелик и составляет (при температуре 370 и парциальном давлении кислорода 100 мм ртутного столба) 0,3 об.%. Это значит, что каждые 100 мл нормальной крови могут переносить в растворенном состоянии лишь 0,3 мл кислорода, что явно недостаточно для поддержания жизнедеятельности организма человека.
В процессе эволюции был выработан принципиально другой механизм переноса кислорода кровью. Этот механизм связан с наличием в крови специального сложного белка (хромопротеида), способного обратимо присоединять молекулярный кислород. В организме человека и высших животных таким хромопротеидом является гемоглобин, содержащийся в эритроцитах.
Под обратимым присоединением кислорода понимают способность гемоглобина образовывать с кислородом лабильное соединение (оксигемоглобин).
Гемоглобин присоединяет кислород в среде с высоким парциальным давлением кислорода и отдает кислород в среде с низким парциальным давлением. Каждый грамм гемоглобина в нормальных условиях может присоединять 1,34 мл кислорода. Поскольку нормальное содержание гемоглобина составляет 14,5 – 16 г.%, то 100 мл крови могут переносить в связанном с гемоглобином состоянии 19 – 21 мл кислорода.
Для того чтобы рассмотреть путь кислорода от легких к тканям, необходимо уточнить вопрос о парциальном давлении кислорода.
Парциальное давление кислорода в воздухе равно 152 мм ртутного столба. Это означает, что из 760 мм нормального атмосферного давления, создаваемого смесью газов воздуха, на долю кислорода приходится приблизительно 1/5 часть. Состав альвеолярного воздуха несколько отличается от состава атмосферного воздуха, главным образом за счет повышения содержания углекислоты. Поэтому содержание кислорода и, следовательно, его парциальное давление в альвеолярном воздухе ниже, чем в атмосферном. Парциальное давление кислорода в альвеолярном воздухе равно 106 – 107 мм ртутного столба. В строгом смысле этого слова понятие «парциальное давление» применимо лишь к смесям газов. В литературе, однако, принято говорить о парциальном давлении кислорода в жидкостях (крови, плазме и т.д.). Под этим подразумевается следующее. Растворимость газа в жидкости зависит от парциального давления данного газа над жидкостью. Для идеальных газов растворимость прямо пропорциональна парциальному давлению газов (закон Генри). Во всяком случае каждому значению содержания газа в жидкости соответствует определенное значение его парциального давления над жидкостью, при котором происходило растворение. Поэтому, когда говорят, например, о том, что парциальное давление кислорода в плазме при каких-то условиях равно 90 мм ртутного столба, это означает, что в плазме в растворенном состоянии содержится столько кислорода, сколько его могло раствориться, если бы плазма находилась в контакте с воздухом, в котором парциальное давление кислорода составляло 90 мм ртутного столба. Аналогичные рассуждения справедливы и для кислорода, связываемого гемоглобином, и для кислорода тканевой жидкости. Это следует иметь в виду при рассмотрении значений парциального давления кислорода в тканях и жидкостях организма.
Парциальное давление кислорода в артериальной крови равно 100-85 мм, парциальное давление кислорода в тканевых жидкостях – 10-20 мм, парциальное давление кислорода в венозной крови – 40-50 мм ртутного столба. Из этих цифр становится ясным, что между тканевой жидкостью и кровью, проходящей по тканевым капиллярам, полное равновесие установиться не успевает.
Рассмотрим теперь, как происходит процесс переноса кислорода. В притекающей к альвеолам венозной крови парциальное давление кислорода составляет около 40 мм ртутного столба. Сквозь мембрану капилляров легочных альвеол кровь соприкасается с кислородом при парциальном давлении 106 мм ртутного столба. Поэтому за время прохождения крови по легочным капиллярам происходит диффузия кислорода в кровь. Следует отметить, что при этом весь кислород должен пройти через стадию растворения в плазме.
Действительно, можно считать, что непосредственно с газообразным кислородом (через мембраны) контактирует плазма. Количество кислорода в плазме венозной крови меньше, чем то количество его, которое может раствориться при парциальном давлении кислорода в альвеолярном воздухе. Поэтому в плазме растворяются дополнительные количества кислорода. В результате нарушается динамическое равновесие между кислородом, растворенным в плазме, и кислородом, связанным гемоглобином в эритроцитах. Кислород из плазмы, диффундируя через мембрану эритроцита, дополнительно связывается с гемоглобином. Если бы этот процесс мог идти достаточно долго, то количество кислорода в оттекающей от альвеол артериальной крови должно было бы соответствовать парциальному давлению в альвеолярном воздухе. Однако за время прохождения крови через альвеолы равновесие установиться полностью не успевает, и парциальное давление кислорода в артериальной крови не превышает 100 мм ртутного столба. Как мы увидим в дальнейшем, этот процесс тесно связан с транспортом углекислоты.
Очевидно, оксигенация крови в альвеолах зависит от большого числа фактороз: скорости кровотока, проницаемости мембраны капилляров и эритроцитов, парциального давления кислорода во вдыхаемом воздухе. В норме толщина мембраны легочной альвеолы равна 4m.
В тканевых капиллярах в процессе отдачи кислорода тканям он также проходит стадию растворения в плазме. С тканевой жидкостью через мембрану контактирует непосредственно плазма крови. В связи с тем что в плазме кислорода содержится больше, чем в тканевой жидкости, диффузия кислорода из крови в ткань превышает диффузию из ткани в кровь, и содержание кислорода в плазме уменьшается. Это приводит к нарушению динамического равновесия между содержанием кислорода в плазме и количеством его, связанным с гемоглобином в эритроцитах.
Равновесие оксигемоглобин – гемоглобин смешается в сторону увеличения количества восстановленного гемоглобина. Таким образом, в результате прохождения крови через тканевые капилляры часть кислорода уходит из крови в ткань.
Разница в содержании кислорода в артериальной и венозной крови носит название артерио-венозного различия и для большинства тканей составляет 5-6 об.%.
Максимальное количество кислорода, которое может связать 100 мл крови за счет как связывания гемоглобином, так и растворения в плазме, равно в норме 21 мл. Эта величина носит название кислородной емкости крови и зависит прежде всего от содержания гемоглобина.
Кроме емкости, показателем дыхательной функции крови является содержание кислорода в крови. Как и емкость, эта величина выражается в объемных процентах и соответствует истинному содержанию кислорода в крови при данных конкретных условиях.
Отношение содержания кислорода к емкости, выраженное в процентах, носит название процента насыщения крови кислородом. Ниже приводятся основные характеристики дыхательной функции крови в норме.
Парциальное давление кислорода в атмосферном воздухе составляет примерно 152 мм ртутного столба, в воздухе легочных альвеол – 106-107 мм, в венозной крови – 50-40 мм и в тканях – 0-20 мм.
Содержание гемоглобина у женщин 14,5 г%, у мужчин – 16 г.%. Один грамм гемоглобина может связать 1,34 мл кислорода. Кислородная емкость составляет 21 об.%. Содержание кислорода в артериальной крови 19 об.%, в венозной крови – 14 об.%. Артерио-венозное различие в содержании кислорода для крови и большинства тканей составляет 5 об.%. Парциальное давление углекислоты в атмосферном воздухе составляет 0,2 мм ртутного столба, в воздухе легочных альвеол – 40 мм, в артериальной крови – 40 мм, в венозной крови – 46 мм и в тканях – 40-60 мм.
Таким образом, весь сложный путь от вдыхаемого воздуха через альвеолярные пространства и кровь к тканям кислорода проходит под знаком постепенного падения парциального давления.
Транспорт углекислоты от тканей к альвеолярному воздуху также совершается при постепенном падении парциального давления углекислоты. Парциальное давление кислорода падает:
- а) при поступлении кислорода из внешнего воздуха в систему альвеол, что объясняется наличием там большого количества углекислоты;
- б) при диффузии кислорода через альвеолярную мембрану в артериальную систему.
Учитывая чрезвычайно малую толщину альвеолярной перегородки, можно было бы ожидать, что парциальное давление кислорода в артериальной крови должно быть таким же, как и в альвеолярном воздухе. Но, так как скорость диффузии кислорода через альвеолярную мембрану составляет только 1/20 – 1/30 скорости диффузии углекислоты, кислород проходит не с такой быстротой, чтобы успело выровняться давление по обе стороны. Это и обусловливает различие в парциальном давлении кислорода между артериальной кровью и альвеолярным воздухом.
Итак, парциальное давление кислорода падает при переходе из артериальной системы в венозную и из капиллярной крови в ткани. В крови парциальное давление кислорода создается количеством его, находящимся в состоянии физического растворения, а также кислородом, связанным с гемоглобином крови, «чудесным веществом» (Баркрофт), обладающим способностью воспринимать из воздуха большие количества кислорода и по мере прохождения через капилляры легко и быстро отдавать его тканям. Тем не менее и кислород, растворенный в плазме, является чрезвычайно важным элементом в кислородном бюджете организма. С одной стороны, он находится почти в полном равновесии с альвеолярным воздухом, с другой – определяет снабжение эритроцитов кислородом. Соотношение давлений кислорода в капиллярной крови и в тканях указывает на дальнейшее его падение при переходе в ткань. В тканях давление кислорода оказывается ниже, а давление СО2 выше, чем в крови.
Способы, позволяющие непосредственно определять давление кислорода в тканях, отсутствуют, поэтому для суждения о нем приходится пользоваться косвенными методами. В настоящее время принято считать, что парциальное давление кислорода в разных тканях различно, будучи обусловлено функциональной способностью ткани. Количество кислорода, доставляемое тканям, значительно превосходит количество, воспринимаемое ими. Ткани обладают специальными механизмами, регулирующими парциальное давление кислорода в них соответственно их потребностям.
Таким образом, переход кислорода из альвеолярного воздуха в кровь есть результат простой диффузии. Давление кислорода в артериальной крови всегда ниже, чем в альвеолярном воздухе. Давление кислорода в тканевой жидкости зависит от количества его, находящегося в растворенном виде, что в свою очередь обусловлено жизнедеятельностью тканей, сопровождающейся постоянным понижением парциального давления кислорода в тканевой жидкости.
Одной из основных функций, которые выполняет кровь в организме животного и человека, является дыхательная функция. Кровь, осуществляя транспорт кислорода воздуха от капилляров легочных альвеол к тканевым капиллярам, обеспечивает таким образом бесперебойную доставку кислорода, необходимого для нормального течения жизненно важных процессов в организме.
В процессе эволюции был выработан принципиально другой механизм переноса кислорода кровью. Этот механизм связан с наличием в крови специального сложного белка (хромопротеида), способного обратимо присоединять молекулярный кислород. В организме человека и высших животных таким хромопротеидом является гемоглобин, содержащийся в эритроцитах.
Под обратимым присоединением кислорода понимают способность гемоглобина образовывать с кислородом лабильное соединение (оксигемоглобин).
Весь сложный путь от вдыхаемого воздуха через альвеолярные пространства и кровь к тканям кислорода проходит под знаком постепенного падения парциального давления.
Парциальное давление кислорода падает при переходе из артериальной системы в венозную и из капиллярной крови в ткани. В крови парциальное давление кислорода создается количеством его, находящимся в состоянии физического растворения, а также кислородом, связанным с гемоглобином крови, «чудесным веществом» (Баркрофт), обладающим способностью воспринимать из воздуха большие количества кислорода и по мере прохождения через капилляры легко и быстро отдавать его тканям. Тем не менее и кислород, растворенный в плазме, является чрезвычайно важным элементом в кислородном бюджете организма. С одной стороны, он находится почти в полном равновесии с альвеолярным воздухом, с другой – определяет снабжение эритроцитов кислородом. Соотношение давлений кислорода в капиллярной крови и в тканях указывает на дальнейшее его падение при переходе в ткань. В тканях давление кислорода оказывается ниже, а давление СО2 выше, чем в крови.
Переход кислорода из альвеолярного воздуха в кровь есть результат простой диффузии. Давление кислорода в артериальной крови всегда ниже, чем в альвеолярном воздухе. Давление кислорода в тканевой жидкости зависит от количества его, находящегося в растворенном виде, что в свою очередь обусловлено жизнедеятельностью тканей, сопровождающейся постоянным понижением парциального давления кислорода в тканевой жидкости.