У молекулы гемоглобина есть только третичная структура

ТЕМА БЕЛКИ
Белки – полимеры, мономерами которых являются аминокислоты.
Среди органических веществ белки занимают первое место по количеству и по значению. В организме человека встречаются 5 млн разнообразных белковых молекул, отличающихся не только друг от друга, но и от белков других организмов. Несмотря на такое разнообразие и сложность строения они построены всего из 20 различных аминокислот.
Строение аминокислоты:
В левой части молекулы расположены группаH2N–, которая обладает свойствами основания; справа – группа –COOH– кислотная, характерная для всех органических кислот. Следовательно, аминокислоты – амфотерные соединения, совмещающие свойства и кислоты и основания. Этим обусловлена их способность взаимодействовать друг с другом. Соединяясь, молекулы аминокислот образуют связи между углеродом кислотной и азотом основной групп. Такие связи называются ковалентными, а в данном случае – пептидными связями:
Соединение двух аминокислот в одну молекулу называется дипептидом, трех аминокислот – трипептидом и т. д., а соединение, состоящее из 20 и более аминокислотных остатков, – полипептидом.
Последовательность аминокислот в полипептидной цепи принято называть первичной структурой белка.
Однако молекула белка в виде цепи аминокислотных остатков, последовательно соединенных между собой пептидными связями, еще не способна выполнять специфические функции. Для этого необходима более высокая структурная организация. Путем образования водородных связей между остатками карбоксильных и аминогрупп разных аминокислот белковая молекула принимает вид спирали (α-структура) или складчатого слоя – «гармошки» (β-структура). Это вторичная структура белка. Но и ее часто недостаточно для приобретения характерной биологической активности.
Часто только молекула, обладающая третичной структурой, может выполнять роль катализатора или любую другую. Третичная структура образуется благодаря взаимодействию радикалов, в частности радикалов аминокислоты цистеина, которые содержат серу. Атомы серы двух аминокислот, находящихся на некотором расстоянии друг от друга в полипептидной цепи, соединяются, образуя так называемые дисульфидные, или S–S, связи. Благодаря этим взаимодействиям, а также другим, менее сильным связям, белковая спираль сворачивается и приобретает форму шарика, или глобулы. Способ укладки полипептидных спиралей в глобуле называют третичной структурой белка. Многие белки, обладающие третичной структурой, могут выполнять свою биологическую роль в клетке. Однако для осуществления некоторых функций организма требуется участие белков с еще более высоким уровнем организации.
Такую организацию называют четвертичной структурой. Присутствует не у всех белков. Она представляет собой функциональное объединение нескольких (двух, трех и более) молекул белка, обладающих третичной структурной организацией. Пример такого сложного белка – гемоглобин. Его молекула состоит из четырех связанных между собой молекул. Другим примером может служить гормон поджелудочной железы – инсулин, включающий два компонента. В состав четвертичной структуры некоторых белков включаются помимо белковых субъединиц и разнообразные небелковые компоненты. Тот же гемоглобин содержит сложное гетероциклическое соединение, в состав которого входит железо.
Строение белковой молекулы:
А – первичная;
Б – вторичная;
В – третичная;
Г – четвертичная структура
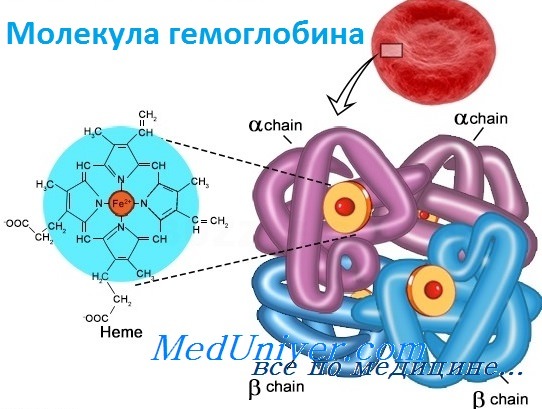
Строение молекулы гемоглобина
Гемоглобин – белок четвертичной структуры. В молекуле гемоглобина белковый компонент представлен белком глобином, небелковый компонент – гем. Глобин состоит из 4 субъединиц. Внутри каждой субъединицы имеется гидрофобный «карман», в котором располагается гем. Содержащийся в геме атом железа связывает кислород.
Свойства белка
Белки, как и другие неорганические и органические соединения, обладают рядом физико-химических свойств:
1. Белки – преимущественно водорастворимые молекулы и, следовательно, могут проявлять свою функциональную активность только в водных растворах.
2. Белковые молекулы несут большой поверхностный заряд. Это определяет целый ряд электрохимических эффектов, например изменение проницаемости мембран каталитической активности и других функций.
3. Белки термолабильны, то есть проявляют свою активность в узких температурных рамках.
Источник
Структура и функция гемоглобинаБолезни гемоглобинов человека, названные гемоглобинопатиями, занимают уникальную позицию в медицинской генетике. Это наиболее частые моногенные болезни у людей, к тому же вызывающие высокую смертность. По оценке Всемирной организации здравоохранения более 5% человеческой популяции — носители мутаций клинически значимых нарушений гемоглобина. Кроме того, поскольку гемоглобин был одной из первых исследованных белковых структур, а гены глобина у человека стали первыми клонированными генами, связанными с болезнями, их молекулярная и биохимическая патология изучена лучше, чем в любой другой группе генетических болезней. Глобины также проливают свет на процесс как молекулярной, так и популяционной эволюции и служат моделью действия генов в развитии. Прежде чем подробно обсуждать гемоглобинопатии, опишем вкратце нормальное состояние гена глобина и биологию гемоглобина. Гемоглобин — переносчик кислорода в эритроцитах позвоночных. Его молекула содержит четыре субъединицы: две а-цепи и две b-цепи. Каждая субъединица формируется цепью полипептида глобина и простетической группой, гемом, железосодержащим пигментом, соединяющимся с кислородом, придавая молекуле способность транспортировать кислород.
Каждая молекула гемоглобина состоит из двух пар полипептидных цепей разного типа. Цепи глобина в нормальном гемоглобине взрослых, гемоглобине А (НЬА), названы а и b соответственно. Четыре цепи складываются и объединяются, формируя шарообразный тетрамер с молекулярным весом приблизительно 64 500, структуру, обозначаемую для НbА — а2b2. Оба типа цепей почти равны по длине; а-цепь имеет 141 аминокислоту, b-цепь — 146. Основные характеристики структуры глобина весьма консервативно сохранялись в ходе эволюции, что стало основой для понимания гемоглобинопатии. Более того, третичная структура полипептида глобина также оказалась в высшей степени консервативной: фактически все изученные глобины имеют семь или восемь извитых участков (в зависимости от цепи). В то же время только две аминокислоты сохраняются во всех глобинах в природе, и неудивительно, что мутации в любом из этих остатков связаны с болезнью. Исследование структуры гемоглобина позволило предсказать, какие типы мутаций, вероятно, окажутся патогенными. Таким образом, мутации, изменяющие строение глобина, замена высококонсервативной аминокислоты или одного из неполярных остатков, формирующих гидрофобную оболочку, исключающую попадание воды внутрь молекулы, вероятно, вызовут гемоглобинопатию. Как и все белки, глобин имеет «чувствительные области», мутации в которых не могут не влиять на функцию, и «нечувствительные области», мутации в которых переносятся более свободно. – Также рекомендуем “Гены гемоглобина человека” Оглавление темы “Выявление генов болезни”:
|
Источник
Гемоглобин входит в состав группы белков гемопротеины, которые сами являются подвидом хромопротеинов и подразделяются на неферментативные белки (гемоглобин, миоглобин) и ферменты (цитохромы, каталаза, пероксидаза). Небелковой частью их является гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
Строение гемоглобина
Строение гемоглобина А
Гемоглобин представляет собой белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями, при этом они взаимодействуют не произвольно, а определенным участком – контактной поверхностью. Этот процесс высокоспецифичен, контакт происходит одновременно в десятках точек по принципу комплементарности. Взаимодействие осуществляют разноименно заряженные группы, гидрофобные участки, неровности на поверхности белка.
Белковые субъединицы в нормальном гемоглобине могут быть представлены различными типами полипептидных цепей: α, β, γ, δ, ε, ξ (соответственно, греч. – альфа, бета, гамма, дельта, эпсилон, кси). В состав молекулы гемоглобина входят по две цепи двух разных типов.
Гем соединяется с белковой субъединицей, во-первых, через остаток гистидина координационной связью железа, во-вторых, через гидрофобные связи пиррольных колец и гидрофобных аминокислот. Гем располагается как бы “в кармане” своей цепи и формируется гемсодержащий протомер.
Нормальные формы гемоглобина
Существует несколько нормальных вариантов гемоглобина:
- HbР (primitive) – примитивный гемоглобин, содержит 2ξ- и 2ε-цепи, встречается в эмбрионе между 7-12 неделями жизни,
- HbF (foetal) – фетальный гемоглобин, содержит 2α- и 2γ-цепи, появляется через 12 недель внутриутробного развития и является основным после 3 месяцев,
- HbA (adult) – гемоглобин взрослых, доля составляет 98%, содержит 2α- и 2β-цепи, у плода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина,
- HbA2 – гемоглобин взрослых, доля составляет 2%, содержит 2α- и 2δ-цепи,
- HbO2 – оксигемоглобин, образуется при связывании кислорода в легких, в легочных венах его 94-98% от всего количества гемоглобина,
- HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
Патологические формы гемоглобина
HbS – гемоглобин серповидно-клеточной анемии.
MetHb – метгемоглобин, форма гемоглобина, включающая трехвалентный ион железа вместо двухвалентного. Такая форма образуется спонтанно, при взаимодействии молекулы O2 и гемового Fe2+, но обычно ферментативных мощностей клетки хватает на его восстановление. При использовании сульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления Fe3+ в Fe2+ в клинике используют аскорбиновую кислоту и метиленовую синь.
Hb-CO – карбоксигемоглобин, образуется при наличии СО (угарный газ) во вдыхаемом воздухе. Он постоянно присутствует в крови в малых концентрациях, но его доля может колебаться от условий и образа жизни.
Угарный газ является активным ингибитором гем-содержащих ферментов, в частности, цитохромоксидазы, 4-го комплекса дыхательной цепи.
Карбоксигемоглобин присутствует и в норме в количестве 0,5-1,5%, в сельской местности меньше, чем в городе. У курильщиков концентрация Hb-CO возрастает, в зависимости от количества сигарет в день, до 8-9%.
HbA1С – гликозилированный гемоглобин. Концентрация его нарастает при хронической гипергликемии и является хорошим скрининговым показателем уровня глюкозы крови за длительный период времени (время жизни эритроцита, 3-4 месяца).
Источник
Белковые молекулы представляют собой линейные гетерополимеры различной длины, мономерами которых являются аминокислоты. В состав белков входят углерод, водород, кислород и азот. Кроме того, почти все они содержат серу, так как она входит в состав аминокислот цистеина и метионина. К некоторым белкам после синтеза могут присоединяться остатки фосфорной кислоты, а также неаминокислотные группы, содержащие железо, магний, цинк, медь, марганец и другие микроэлементы. Такие белки называют сложными. В них выделяют полипептидную часть и небелковую часть, или простетическую группу. Так, например, в состав белка гемоглобина входит полипептид глобин и небелковая группа — гем, содержащая ион железа. Среди сложных белков в зависимости от природы простетической группы выделяют:
хромопротеины (содержат пигменты);
металлопротеины (содержат металлы);
липопротеины (содержат липиды);
нуклеопротеины (содержат нуклеиновые кислоты) и др.
Пептид, содержащий более 40–50 аминокислотных остатков, обычно называют полипептидом или белком. Таким образом, разница между белком и пептидом заключается в том, что пептидом обычно называют низкомолекулярное соединение, а белком — высокомолекулярное. Молекулы белка могут содержать сотни и даже тысячи аминокислотных остатков: молекулярная масса белков колеблется в пределах от нескольких тысяч до сотен тысяч и даже миллионов дальтон.
Первичная структура белка
Каждая белковая молекула в живом организме характеризуется определенной последовательностью аминокислот, которая задается последовательностью нуклеотидов в структуре гена, кодирующего данный белок. Таким образом, в организме синтезируются белки с точно определенной химической структурой, которые были отобраны для выполнения определенных функций в процессе эволюции.
Последовательность аминокислотных остатков в молекуле белка определяет его первичную структуру, то есть его химическую формулу. Точно так же как алфавит, в состав которого входят 33 буквы, позволяет создать огромное количество слов, с помощью 20 аминокислот можно создать почти неограниченное количество разнообразных белков. Общее число белков с различной первичной структурой, встречающихся в организме человека, оценивается примерно в 50 000, а у всех видов живых организмов составляет величину порядка $mathrm{10^{10}-10^{12}}$.
Аминокислотные остатки в белке связаны между собой пептидной связью.

Рис. 1
Пептидная связь имеет ряд особенностей, которые в значительной степени влияют на укладку полипептидной цепи в пространстве (см. рис. 1). В результате взаимодействия неподеленных электронов атома азота, участвующего в образовании пептидной связи, с электронами, образующими расположенную рядом двойную связь С=О, происходит перераспределение электронной плотности и изменение свойств пептидной связи (С–N). Она приобретает характер двойной связи.
Благодаря особенностям пептидной связи шесть атомов пептидной группировки (атомы С и N, участвующие в образовании пептидной связи, а также кислород и водород, присоединенные к этим атомам, и два С$alpha$-атома соседних аминокислот) оказываются расположенными в одной плоскости. Таким образом, полипептидную цепь можно представить как набор относительно жестких плоских структур, имеющих общую точку вращения при С$alpha$-атоме.
Пептидная связь достаточно прочна, ее расщепление происходит лишь при использовании химических катализаторов (кислота или основание) в жестких условиях (например, инкубации в течение 24 часов в 6 н HCl при температуре 105 оС), либо при катализе специфическими ферментами — пептидазами.
В пептидной или белковой цепи выделяют N-концевой остаток, содержащий свободную аминогруппу, и С-концевой остаток, содержащий карбоксильную группу. Последовательность аминокислот в полипептидной цепи записывается, начиная с N-конца. Для обозначения аминокислот в полипептидной цепи существует трехбуквенный и однобуквенные коды аминокислот. В соответствии с трехбуквенным кодом последовательность аминокислот в пятичленном пептиде аланин-гистидин-глицин-цистеин-лейцин записывается как Аlа-His-Gly-Cys-Leu.
Вторичной структурой белка называют пространственное расположение полипептидной цепи белка на отдельных ее участках в виде спирали или слоя (листа). Пептидные связи полипептидной цепи содержат группы С=О и N–H, способные образовывать внутримолекулярные водородные связи — основной тип нековалентных взаимодействий, стабилизирующий элементы вторичной структуры.
Главную роль в стабилизации периодически повторяющихся элементов вторичной структуры белка играют водородные связи между пептидными группами (C=O…H–N). Основными элементами вторичной структуры, встречающимися в различных белках, являются $alpha$-спираль и $beta$-складчатый слой (или лист), хотя встречаются и другие элементы вторичной структуры.
типы вторичных структур белка
альфа-спирали

Рис. 2
При формировании $alpha$-спирали происходит закручивание скелета полипептидной цепи в виде спирали вокруг воображаемой оси, сопровождающееся сближением пептидных связей, что благоприятствует образованию внутримолекулярных водородных связей между NH-группами, находящимися на одном витке, и CO-группами, расположенными на другом витке спирали. Направление этих связей параллельно оси спирали (см. рис. 2). Боковые цепи аминокислот располагаются с наружной стороны спирали. Длина витка $alpha$-спирали составляет 5,4 ангстремы, в одном таком витке укладываются 3,6 аминокислотных остатка. В белках встречается только правозакрученная $alpha$-спираль. $alpha$-спираль стабилизируется не только водородными связями, но и за счет взаимодействия боковых радикалов, располагающихся друг над другом в соседних витках спирали. По этой причине появление в первичной последовательности разноименно заряженных или гидрофобных аминокислот через 3–4 аминокислотных остатка будет стабилизировать $alpha$-спираль, и напротив, наличие одноименно заряженных аминокислот через 3–4 аминокислотных остатка приведет к их отталкиванию, что не позволит образоваться $alpha$-спирали.
бета-слои

Рис. 3
Второй элемент вторичной структуры, который часто встречается в белках, называется $beta$-слоем (листом), или $beta$-складчатым слоем (листом) (см. рис. 3). Структура типа складчатого слоя формируется двумя параллельно или антипараллельно расположенными участками полипептидной цепи. Она стабилизируется за счет водородных связей, которые образуются между расположенными рядом остовами полипептидной цепи. Боковые цепи аминокислот располагаются перпендикулярно по отношению к плоскости остова цепи, который при этом изгибается, образуя плоский лист. Хотя водородные связи слабее ковалентных, присутствие их в значительном количестве делает структуры типа a-спирали или b-складчатого слоя достаточно прочными. Существуют два типа $beta$-слоев — параллельный и антипараллельный, в зависимости от того, меняется ли направление хода полипептидной цепи в каждом следующем фрагменте, участвующем в образовании слоя.
Также как особый тип $beta$-структур иногда выделяют $beta$-шпильки, представляющие собой два антипараллельно расположенных участка одной и той же цепи, которая разворачивается на вершине шпильки на $180^circ$.
третичная структура белка

Рис. 4
$alpha$-cпиральные участки и $beta$-структуры типа складчатого слоя подвергаются дальнейшей упаковке, в результате чего формируется третичная структура белка. Если вторичная структура характеризует укладку какого-либо участка полипептидной цепи, то третичная структура — это структура всей полипептидной цепи в целом. Растворимые белки обычно бывают глобулярными (от лат. «глобула» — шарик), то есть их молекулы имеют округлые или эллиптические очертания, хотя среди них встречаются и фибриллярные белки — имеющие форму волокна, или фибриллы (от лат. «фибрилла» — волоконце), возможно также сочетание глобулярной и фибриллярной структур. В белковой глобуле заряженные и полярные аминокислотные остатки оказываются на поверхности, а гидрофобные — внутри. $beta$-складчатые слои часто входят в состав сердцевины многих глобулярных белков. В упакованной в виде глобулы молекуле белка зачастую сближаются аминокислотные остатки, которые в полипептидной цепи расположены далеко друг от друга (см. рис.). Нерастворимые в воде белки часто бывают фибриллярными. Примеры третичных структур разных белков можно увидеть на рис. 4.
В принципе, белковая молекула может укладываться различными способами, принимая большое число различных форм (конформаций) в зависимости от условий (рН, температура, наличие ионов). Однако в клетке большинство белков в нативном (неповрежденном) состоянии существует лишь в одной или нескольких близких конформациях, характерных для данного полипептида. Она определяется тем, как сворачивается полипептидная цепь в растворе, что, в свою очередь, зависит от последовательности аминокислот в этой цепи и условий (температура, рН, наличие ионов и т. д). Боковые группы аминокислот взаимодействуют друг с другом и с водой с образованием слабых нековалентных связей (водородных, ионных, гидрофобных).
В некоторых случаях для обеспечения большей стабильности третичной структуры происходит образование ковалентных связей. Это в основном происходит при взаимодействии оказавшихся близко друг к другу SH-групп остатков цистеина, которые окисляются, формируя S–S-связи, или дисульфидные мостики (рис. 5). Образование таких связей особенно характерно для белков, выделяемых из клетки наружу или находящихся в плазматической мембране с наружной стороны, поскольку эти белки оказываются в условиях, значительно отличающихся от тех, что существуют внутри клетки.

Рис. 5
четвертичная структура белка
Многие белки состоят из нескольких полипептидных цепей одинаковой или различной структуры. Объединение белков становится возможным в том случае, если на поверхности белка образуется центр связывания для того же самого или другого белка. При объединении нескольких полипептидных цепей образуется белок, для которого характерна четвертичная структура. Такие белки называют олигомерами, а входящие в состав олигомера отдельные полипептидные цепи — мономерами, или
субъединицами. Типичным примером олигомерного белка является молекула гемоглобина (рис. 6), состоящая из двух $alpha$-цепей и двух $beta$-цепей.


Рис. 6
Каждая пара полипептидных цепей в молекуле гемоглобина представлена одинаковыми молекулами ($alpha$- и $beta$-глобинами).
Многие олигомерные белки, в свою очередь, являются компонентами, участвующими в формировании более крупных агрегатов.
Таким образом, благодаря определенной последовательности аминокислот в полипептидной цепи отдельные ее части сворачиваются в $alpha$-спирали или формируют $beta$-складки, которые соединяются гибкими участками. Эти элементы вторичной структуры укладываются в пространстве, образуя глобулы или фибриллы, то есть формируют третичную структуру белка. И наконец, отдельные глобулы или фибриллы взаимодействуют друг с другом с образованием комплексов, состоящих из нескольких молекул, что приводит к формированию четвертичной структуры.
Денатурация и ренатурация белков
Большая часть белковых молекул способна сохранять свою биологическую активность, то есть выполнять свойственную им функцию только в узком диапазоне температур и кислотности среды. При повышении температуры, изменении кислотности до экстремальных значений, добавлении гидрофобных агентов (например, органических растворителей) или при значительном увеличении концентрации солей в структуре белков происходят изменения, которые приводят к их денатурации — потере белком своей нативной (естественной) пространственной структуры. Как правило, при этом первичная структура белка не разрушается. Примером денатурации является свертывание белка яйца, наблюдающееся при его варке. Денатурация — это разрушение характерной для данного белка четвертичной, третичной и вторичной структуры, в результате чего в денатурированном состоянии полипептидные цепи белков образуют случайные и беспорядочные клубки и петли. Разрыва пептидных связей при денатурации не происходит, то есть сама полипептидная цепь сохраняется, однако способ ее укладки изменяется. В том случае если в белке имеются дисульфидные мостики, стабилизирующие третичную структуру белка, обычно при денатурации они рвутся, что происходит путем восстановления остатков цистеина.
Денатурация бывает обратимой и необратимой. В случае обратимой денатурации при возвращении в исходные (нативные) условия пространственная структура белка восстанавливается. При варке яйца мы имеем дело с необратимой денатурацией, когда исходную (нативную) структуру восстановить уже практически невозможно. Как правило, необратимая денатурация связана не с нарушением первичной структуры, а с тем, что разные полипептидные цепи взаимодействуют своими гидрофобными участками, слипаются и образуют большие агрегаты — твердые частицы белка, выпадающие в осадок. Свет рассеивается на границе этих частиц, поэтому прозрачный раствор белка (например, белок яйца) становится непрозрачной взвесью твердых частиц белкового агрегата, что объясняет белый цвет и непрозрачность белка вареного яйца.
В клетке также происходит ренатурация белков, обычно поврежденных, отслуживших свой срок. Такие белки либо разрушаются (деградируют), либо, если это еще возможно, ренатурируют — самостоятельно или при помощи белков-шаперонов, своеобразных помощников, способствующих восстановлению структуры других белков. Шапероны играют большую роль в восстановлении клетки после теплового шока.
Рентгеноструктурный анализ
Основным источником знаний о структуре белков является метод рентгеноструктурного анализа. Для того чтобы провести рентгеноструктурный анализ, необходимо получить кристаллы белка, что далеко не всегда удается. Иногда фермент кокристаллизуют (совместно кристаллизуют) с субстратом или ингибитором, другие белки тоже могут кокристаллизоваться с какими-либо веществами. После получения кристаллов белка их облучают рентгеновскими лучами и получают картину дифракции этих лучей на кристаллической структуре белка.

Рис. 7 По положению пятен на дифракционной картине рассчитывают положение каждого атома в молекуле белка. В наилучшем случае удается добиться разрешения 2–3 ангстрем и даже 1 ангстрем (ангстрем = 0,1 нм = $10^{-10}$ м).

Рис. 8

Рис. 9

Рис. 10
Источник