Уровни организации белка гемоглобина

Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
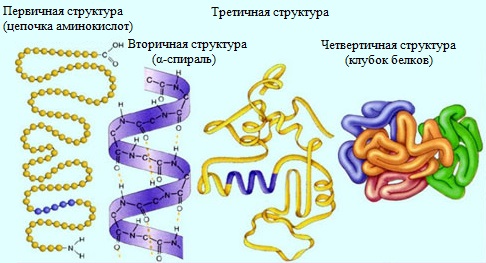
Первичная структура белка – определенная последовательность a-аминокислотных остатков в полипептидной цепи.
Пептидная цепь имеет линейную структуру только у небольшого числа белков. В большинстве белков пептидная цепь определенным образом свернута в пространстве.
Один из первых белков, первичная структура которого была установлена в 1954 г. — гормон инсулин (регулирует содержание сахара в крови), его молекула состоит из двух полипептидных цепей, которые связаны друг с другом (в одной цепи 21 аминокислотный остаток, в другой – 30).
Вторичная структура белка – конформация полипептидной цепи, закрепленная множеством водородных связей между группами N-H и С=О.
В результате образования внутримолекулярных водородных связей между атомами водорода аминогрупп и атомами кислорода карбонильных групп полипептидные цепи многих белков скручиваются в спираль.
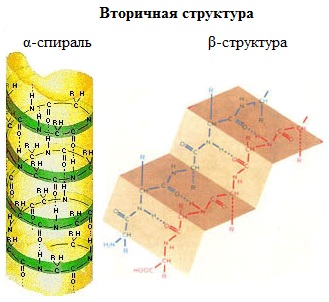
Существует два основных способа укладки цепи.
Одна из моделей вторичной структуры – a-спираль. Другая модель – β-форма («складчатый лист»), в которой преобладают межцепные (межмолекулярные) Н-связи.
В α-спирали на одном витке укладываются четыре аминокислотных остатка. Все радикалы аминокислот находятся снаружи спирали. Между группами NH и СО, находящимися на соседних витках, образуются водородные связи, которые стабилизируют спираль.
В β-структуре (складчатом слое) полипептидная цепь растянута, ее участки располагаются параллельно друг другу и удерживаются водородными связями.
Большинство белков содержит как α-спирали, так и β-структуры.
Вторичная структура была установлена американским химиком Л. Полингом в 1951 г.
Третичная структура белка — форма закрученной спирали в пространстве, образованная главным образом за счет дисульфидных мостиков -S-S- , водородных связей, гидрофобных и ионных взаимодействий.
Третичная структура – это трехмерная пространственная конфигурация закрученной α-спирали или β-структуры в пространстве.
У большинства белков полипептидные цепи свернуты особым образом в «клубок» — компактную «глобулу».
Белок в водном растворе свертывается таким образом, чтобы его гидрофобные (водоотталкивающие — от греч. гидро – вода, фобос – страх) боковые цепи были внутри молекулы, а гидрофильные (растворимые) – повернуты наружу.
Третичная структура образуется за счет дисульфидных мостиков -S-S- между цистеиновыми остатками, находящимися в разных местах полипептидной цепи.
В образовании третичной структуры участвуют также ионные взаимодействия противоположно заряженных групп (солевые мостики) NH3+ и COO— .

Интересно знать!
В составе волос содержится белок кератин. В его молекуле имеется большое количество дисульфидных связей. С помощью химической завивки волосам можно придать другую форму. Для этого волосы сначала накручивают на бигуди, затем обрабатывают раствором реагента-восстановителя, разрушающего дисульфидные связи, и прогревают. В результате этого кератин приобретает иную пространственную структуру. Далее волосы промывают и обрабатывают реагентом-окислителем, при этом происходит образование новых дисульфидных связей. Вследствие этого вновь приобретенная структура кератина стабилизируется. Волосы приобретают другую форму.
Четвертичная структура белка — агрегаты нескольких белковых макромолекул (белковые комплексы), образованные за счет взаимодействия разных полипептидных цепей.
Четвертичная структура – способ совместной укладки нескольких полипептидных цепей. Образующиеся структуры называются ассоциатами.
Термин «четвертичная структура» был предложен в 1958 г. Дж. Берналом.
Характерной особенностью белков с четвертичной структурой является их способность к самосборке, например, гемоглобин (белок крови) легко собирается из смеси α- и β-цепей и гема.

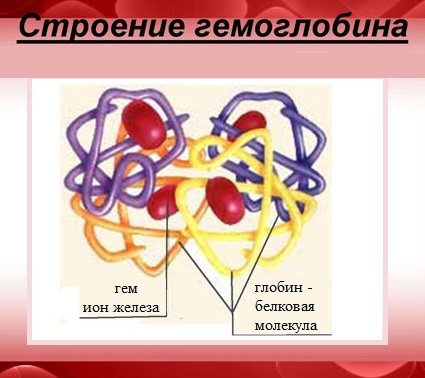
Гемоглобин — сложный белок, макромолекула которого состоит из четырех полипептидных цепей (глобул), соединенных с четырьмя гемами – небелковыми образованиями, которые и придают крови красный цвет.
В каждом геме содержится один атом двухвалентного железа, который может непрочно связывать одну молекулу кислорода. В результате такого связывания образуется оксигемоглобин, одна молекула которого переносит к тканям четыре молекулы кислорода.
Из тканей гемоглобин выносит углекислый газ, молекулы которого присоединяются к аминогруппам, содержащимся в полипептидных цепях.
Белки
Молекула гемоглобина: 4 субъединицы окрашены в разные цвета
Структура гемоглобина человека. Железосодержащие гем-группы показаны зелёным. Красным и синим показаны альфа- и бета- субъединицы.
Гемоглоби́н (от др.-греч. αἷμα «кровь» + лат. globus «шар») (Hb или Hgb) — сложный железосодержащий белок животных, обладающих кровообращением, способный обратимо связываться с кислородом, обеспечивая его перенос в ткани. У позвоночных животных содержится в эритроцитах, у большинства беспозвоночных растворён в плазме крови (эритрокруорин) и может присутствовать в других тканях[1]. Молекулярная масса гемоглобина человека — около 66,8 кДа. Молекула гемоглобина может нести до четырёх молекул кислорода. Один грамм гемоглобина может переносить до 1.34 мл. O2
Гемоглобин появился более чем 400 миллионов лет назад у последнего общего предка человека и акул в результате 2 мутаций, приведших к формированию четырёхкомпонентного комплекса гемоглобина, сродство которого к кислороду достаточно для связывания кислорода в насыщенной им среде, но недостаточно, чтобы удерживать его в других тканях организма.[2][3]
Большой вклад в исследование структуры и функционирования гемоглобина внёс Макс Фердинанд Перуц, получивший за это в 1962 году Нобелевскую премию[4].
Нормальным содержанием гемоглобина в крови человека считается: у мужчин — 130—160 г/л (нижний предел — 120, верхний предел — 180 г/л), у женщин — 120—160 г/л; у детей нормальный уровень гемоглобина зависит от возраста и подвержен значительным колебаниям. Так, у детей через 1—3 дня после рождения нормальный уровень гемоглобина максимален и составляет 145—225 г/л, а к 3—6 месяцам снижается до минимального уровня — 95—135 г/л, затем с 1 года до 18 лет отмечается постепенное увеличение нормального уровня гемоглобина в крови[5].
Во время беременности в организме женщины происходит задержка и накопление жидкости, что является причиной гемодилюции — физиологического разведения крови. В результате наблюдается относительное снижение концентрации гемоглобина (при беременности уровень гемоглобина в норме составляет 110—155 г/л). Кроме этого, в связи с внутриутробным ростом ребёнка происходит быстрое расходование запасов железа и фолиевой кислоты. Если до беременности у женщины был дефицит этих веществ, проблемы, связанные со снижением гемоглобина, могут возникнуть уже на ранних сроках беременности[6].
Главные функции гемоглобина: перенос кислорода и буферная функция. У человека в капиллярах лёгких в условиях избытка кислорода последний соединяется с гемоглобином. Потоком крови эритроциты, содержащие молекулы гемоглобина со связанным кислородом, доставляются к органам и тканям, где кислорода мало; здесь необходимый для протекания окислительных процессов кислород освобождается от связи с гемоглобином. Кроме того, гемоглобин способен связывать в тканях небольшое количество диоксида углерода (CO2) и освобождать его в лёгких.
Монооксид углерода (CO) связывается с гемоглобином крови намного сильнее (в 250 раз[7]), чем кислород, образуя карбоксигемоглобин (HbCO). Впрочем, монооксид углерода может быть частично вытеснен из гема при повышении парциального давления кислорода в лёгких. Некоторые процессы приводят к окислению иона железа в гемоглобине до степени окисления +3. В результате образуется форма гемоглобина, известная как метгемоглобин (HbOH) (metHb, от «мета-» и «гемоглобин», иначе гемиглобин или ферригемоглобин, см. Метгемоглобинемия). В обоих случаях блокируются процессы транспортировки кислорода.
Строение[править | править код]
Гемоглобин является сложным белком класса гемопротеинов, то есть в качестве простетической группы здесь выступает гем — порфириновое ядро, содержащее железо. Гемоглобин человека является тетрамером, то есть состоит из 4 протомеров. У взрослого человека они представлены полипептидными цепями α1, α2, β1 и β2. Субъединицы соединены друг с другом по принципу изологического тетраэдра. Основной вклад во взаимодействие субъединиц вносят гидрофобные взаимодействия. И α-, и β-цепи относятся к α-спиральному структурному классу, так как содержат исключительно α-спирали. Каждая цепь содержит восемь спиральных участков, обозначаемых буквами от A до H (от N-конца к C-концу).
Гем представляет собой комплекс протопорфирина IX, относящегося к классу порфириновых соединений, с атомом железа(II). Этот кофактор нековалентно связан с гидрофобной впадиной молекул гемоглобина и миоглобина.
Железо(II) характеризуется октаэдрической координацией, то есть связывается с шестью лигандами. Четыре из них представлены атомами азота порфиринового кольца, лежащими в одной плоскости. Две другие координационные позиции лежат на оси, перпендикулярной плоскости порфирина. Одна из них занята азотом остатка гистидина в 93-м положении полипептидной цепи (участок F). Связываемая гемоглобином молекула кислорода координируется к железу с обратной стороны и оказывается заключённой между атомом железа и азотом ещё одного остатка гистидина, располагающегося в 64-м положении цепи (участок E).
Всего в гемоглобине человека четыре участка связывания кислорода (по одному гему на каждую субъединицу), то есть одновременно может связываться четыре молекулы. Гемоглобин в лёгких при высоком парциальном давлении кислорода соединяется с ним, образуя оксигемоглобин. При этом кислород соединяется с гемом, присоединяясь к железу гема на 6-ю координационную связь. На эту же связь присоединяется и монооксид углерода, вступая с кислородом в «конкурентную борьбу» за связь с гемоглобином, образуя карбоксигемоглобин.
Связь гемоглобина с монооксидом углерода более прочная, чем с кислородом. Поэтому часть гемоглобина, образующая комплекс с монооксидом углерода, не участвует в транспорте кислорода. В норме у человека образуется 1,2 % карбоксигемоглобина. Повышение его уровня характерно для гемолитических процессов, в связи с этим уровень карбоксигемоглобина является показателем гемолиза.
Физиология[править | править код]
Изменение состояний окси- и дезоксигемоглобина
В отличие от миоглобина гемоглобин имеет четвертичную структуру, которая придаёт ему способность регулировать присоединение и отщепление кислорода и характерную кооперативность: после присоединения первой молекулы кислорода связывание последующих облегчается. Структура может находиться в двух устойчивых состояниях (конформациях): оксигемоглобин (содержит 4 молекулы кислорода; напряжённая конформация) и дезоксигемоглобин (кислорода не содержит; расслабленная конформация).
Устойчивое состояние структуры дезоксигемоглобина усложняет присоединение к нему кислорода. Поэтому для начала реакции необходимо достаточное парциальное давление кислорода, что возможно в альвеолах лёгких. Изменения в одной из 4-х субъединиц влияет на оставшиеся, и после присоединения первой молекулы кислорода связывание последующих облегчается.
Отдав кислород тканям, гемоглобин присоединяет к себе ионы водорода и углекислый газ, перенося их в лёгкие[8].
Гемоглобин является одним из основных белков, которыми питаются малярийные плазмодии — возбудители малярии, и в эндемичных по малярии районах земного шара весьма распространены наследственные аномалии строения гемоглобина, затрудняющие малярийным плазмодиям питание этим белком и проникновение в эритроцит. В частности, к таким имеющим эволюционно-приспособительное значение мутациям относится аномалия гемоглобина, приводящая к серповидноклеточной анемии. Однако, к несчастью, эти аномалии (как и аномалии строения гемоглобина, не имеющие явно приспособительного значения) сопровождаются нарушением кислород-транспортирующей функции гемоглобина, снижением устойчивости эритроцитов к разрушению, анемией и другими негативными последствиями. Аномалии строения гемоглобина называются гемоглобинопатиями.
Гемоглобин высокотоксичен при попадании значительного его количества из эритроцитов в плазму крови (что происходит при массивном внутрисосудистом гемолизе, геморрагическом шоке, гемолитических анемиях, переливании несовместимой крови и других патологических состояниях). Токсичность гемоглобина, находящегося вне эритроцитов, в свободном состоянии в плазме крови, проявляется тканевой гипоксией — ухудшением кислородного снабжения тканей, перегрузкой организма продуктами разрушения гемоглобина — железом, билирубином, порфиринами с развитием желтухи или острой порфирии, закупоркой почечных канальцев крупными молекулами гемоглобина с развитием некроза почечных канальцев и острой почечной недостаточности.
Ввиду высокой токсичности свободного гемоглобина в организме существуют специальные системы для его связывания и обезвреживания. В частности, одним из компонентов системы обезвреживания гемоглобина является особый плазменный белок гаптоглобин, специфически связывающий свободный глобин и глобин в составе гемоглобина. Комплекс гаптоглобина и глобина (или гемоглобина) затем захватывается селезёнкой и макрофагами тканевой ретикуло-эндотелиальной системы и обезвреживается.
Другой частью гемоглобинообезвреживающей системы является белок гемопексин[en], специфически связывающий свободный гем и гем в составе гемоглобина. Комплекс гема (или гемоглобина) и гемопексина затем захватывается печенью, гем отщепляется и используется для синтеза билирубина и других жёлчных пигментов, или выпускается в рециркуляцию в комплексе с трансферринами для повторного использования костным мозгом в процессе эритропоэза.
Экспрессия генов гемоглобина до и после рождения.
Также указаны типы клеток и органы, в которых происходит экспрессия гена (данные по Wood W. G., (1976). Br. Med. Bull. 32, 282.).[9]
Гемоглобин при заболеваниях крови[править | править код]
Дефицит гемоглобина может быть вызван, во-первых, уменьшением количества молекул самого гемоглобина (см. анемия), во-вторых, из-за уменьшенной способности каждой молекулы связать кислород при том же самом парциальном давлении кислорода.
Гипоксемия — это уменьшение парциального давления кислорода в крови, её следует отличать от дефицита гемоглобина. Хотя и гипоксемия, и дефицит гемоглобина являются причинами гипоксии.
Если дефицит кислорода в организме в общем называют гипоксией, то местные нарушения кислородоснабжения называют ишемией.
Прочие причины низкого гемоглобина разнообразны: кровопотеря, пищевой дефицит, болезни костного мозга, химиотерапия, отказ почек, атипичный гемоглобин.
Повышенное содержание гемоглобина в крови связано с увеличением количества или размеров эритроцитов, что наблюдается также при истинной полицитемии. Это повышение может быть вызвано: врождённой болезнью сердца, лёгочным фиброзом, слишком большим количеством эритропоэтина.
См. также[править | править код]
- Гемоглобин А
- Гемоглобин С (мутантная форма)
- Эмбриональный Гемоглобин (эмбриональный)
- Гемоглобин S (мутантная форма)
- Гемоглобин F (фетальный)
- Кобоглобин
- Нейроглобин
- Анемия
- Порфирия
- Талассемия
- Эффект Вериго — Бора
Примечания[править | править код]
- ↑ Haemoglobins of invertebrate tissues. Nerve haemoglobins of Aphrodite, Aplysia and Halosydna
- ↑ Ученые выяснили происхождение гемоглобина. РИА Новостей, 20.05.2020, 18:59
- ↑ Michael Berenbrink. Evolution of a molecular machine/Nature, NEWS AND VIEWS, 20 MAY 2020
- ↑ Лауреаты нобелевской премии. Макс Перуц.
- ↑ Назаренко Г. И., Кишкун А. А. Клиническая оценка результатов лабораторных исследований. — 2005.
- ↑ Общий анализ крови и беременность Архивная копия от 10 марта 2014 на Wayback Machine
- ↑ Hall, John E. Guyton and Hall textbook of medical physiology (англ.). — 12th ed.. — Philadelphia, Pa.: Saunders/Elsevier, 2010. — P. 1120. — ISBN 978-1416045748.
- ↑ Степанов В. М. Структура и функции белков : Учебник. — М. : Высшая школа, 1996. — С. 167—175. — 335 с. — 5000 экз. — ISBN 5-06-002573-X.
- ↑ Айала Ф., . Современная генетика: В 3-х т = Modern Genetics / Пер. А. Г. Имашевой, А. Л. Остермана, . Под ред. Е. В. Ананьева. — М.: Мир, 1987. — Т. 2. — 368 с. — 15 000 экз. — ISBN 5-03-000495-5.
Литература[править | править код]
- Mathews, CK; KE van Holde & KG Ahern (2000), Biochemistry (3rd ed.), Addison Wesley Longman, ISBN 0-8053-3066-6
- Levitt, M & C Chothia (1976), “Structural patterns in globular proteins”, Nature
Ссылки[править | править код]
- Eshaghian, S; Horwich, TB; Fonarow, GC (2006). “An unexpected inverse relationship between HbA1c levels and mortality in patients with diabetes and advanced systolic heart failure”. Am Heart J. 151 (1): 91.e1—91.e6. DOI:10.1016/j.ahj.2005.10.008. PMID 16368297.
- Kneipp J, Balakrishnan G, Chen R, Shen TJ, Sahu SC, Ho NT, Giovannelli JL, Simplaceanu V, Ho C, Spiro T (2005). “Dynamics of allostery in hemoglobin: roles of the penultimate tyrosine H bonds”. J Mol Biol. 356 (2): 335—53. DOI:10.1016/j.jmb.2005.11.006. PMID 16368110.
- Hardison, Ross C. (2012). “Evolution of Hemoglobin and Its Genes”. Cold Spring Harbor Perspectives in Medicine. 2 (12): a011627. DOI:10.1101/cshperspect.a011627. ISSN 2157-1422. PMC 3543078. PMID 23209182.
Патология белкового обмена
Белки – неразветвляющиеся полимеры, минимальная структурная единица которых – аминокислота (АК).
Аминокислоты соединены между собой пептидной связью. В состав белков входит 20 АК в альфа-форме, расположенных в различной, но строго определенной для каждого белка последовательности.
Белки организма включают около 16% азота, который в ряде методов является маркером вещества белкового происхождения.
Сведения о суммарном количестве белков плазмы (общий белок крови) получают обычно рефрактометрическим и фотометрическим биуретовыми методами.
Плазма крови человека в норме содержит более 100 видов белков.
Около 20% общего белка составляют альбумины, иммуноглобулины, липопротеиды, фибриноген, трансферрин и др.
Нормальные величины общего белка плазмы: 65-85 г/л.
Классификация аминокислот
I. По химическому строению
§ Алифатические – глицин (Гли), аланин (Ала), валин (Вал), лейцин (Лей), изолейцин (Илей);
§ Оксикислоты – серин (Сер), треанин (Тре);
§ Дикарбоновые – аспарагин (Асп), глутамин (Глу), аспарагиновая кислота (Аск), глутаминовая кислота (Глк);
§ Двуосновные – лизин (Лиз), гистидин (Гис), аргинин (Арг);
§ Ароматические – фениналанин (Фен), тирозин (Тир), триптофан (Три);
§ Серосодержащие – цистеин (Цис), метионин (Мет).
II. По биохимической роли:
§ глюкогенные – через ряд химических превращений поступают на путь гликолиза (окисления глюкозы) – Гли, Ала, Тре, Вал, Аск, Глк, Арг, Гис, Мет.
§ кетогенные – участвуют в образовании кетоновых тел – Лей, Илей, Тир, Фен.
III. По заменимости:
§ Незаменимые – не синтезируются в организме – Гис, Иле, Лей, Лиз, Мет, Фен, Тре, Три, Вал.
§ Заменимые – остальные.
За счет наличия в молекуле АК одновременно аминной и карбоксильной групп этим соединениям присущи кислотно-основные свойства. В нейтральной среде АК существуют в виде биполярных ионов – цвиттер-ионов т.е. не NH2– –COOH , а NH3+–R-COO – .
Если карбоксильная группа одной АК ацилирует аминогруппу другой АК, то образуется ковалентная амидная связь, которую называют пептидной. Т.о. пептиды – это соединения, образованные из остатков альфа-АК, соединенных между собой пептидной связью.
Рис. 16.1. Образование полипептида из аминокислот с помощью ковалентных пептидных (амидных) связей [https://www.nsu.ru/education/biology/molbiol/Lecture3/Lec32.htm].
Данная связь достаточно стабильна и разрыв ее происходит лишь при участии катализаторов – специфических ферментов. Посредством такой связи АК объединяются в достаточно длинные цепочки, которые носят название полипептидных. Каждая такая цепь содержит на одном конце АК со свободной аминогруппой – это N-концевой остаток, и на другом с карбоксильной группой – С-концевой остаток.
Полипептиды, способные самопроизвольно формировать и удерживать определенную пространственную структуру, которая называется конформацией, относят к белкам.
Стабилизация такой структуры возможна лишь при достижении полипептидами определенной длины, поэтому белками обычно считают полипептиды молекулярной массой более 5 000 Да. (1 Да равен 1/12 изотопа углерода). Только имея определенное пространственное строение, белок может функционировать.
Функции белков
1. Структурная (пластическая) – белками образованы многие клеточные компоненты, а в комплексе с липидами они входят в состав клеточных мембран.
2. Каталитическая – все биологические катализаторы – ферменты по своей химической природе являются белками.
3. Транспортная – белок гемоглобин транспортирует кислород, ряд других белков образуя комплекс с липидами транспортируют их по крови и лимфе (например, миоглобин, сывороточный альбумин).
4. Механохимическая – мышечная работа и иные формы движения в организме осуществляются при непосредственном участии сократительных белков с использованием энергии макроэргических связей (например, актин, миозин).
5. Регуляторная – ряд гормонов и других биологически активных веществ имеют белковую природу (например, инсулин, АКТГ).
6. Защитная – антитела (иммуноглобулины) являются белками, кроме того, основу кожи составляет белок коллаген, а волос – кератин. Кожа и волосы защищают внутреннюю среду организма от внешних воздействий. В состав слизи и синовиальной жидкости входят мукопротеиды.
7. Опорная – сухожилия, поверхности суставов соединения костей образованы в значительной степени белковыми веществами (например, коллаген, эластин).
8. Энергетическая – аминокислоты белков могут поступать на путь гликолиза, который обеспечивает клетку энергией.
9. Рецепторная – многие белки участвуют в процессах избирательного узнавания (рецепторы).
Уровни организации белковой молекулы
В современной литературе принято рассматривать 4 уровня организации структуры молекулы белка.
Последовательность аминокислотных остатков, соединенных между собой пептидной связью называют первичным уровнем организации белковой молекулы. Она кодируется структурным геном каждого белка. Связи: пептидная и дисульфидные мостики между относительно близко расположенными остатками цистеинов. Это ковалентные взаимодействия, которые разрушаются только под действием протеолитических ферментов (пепсин, трипсин и т.д.).
Рис. 16.2. Первичная (А) и вторичная (В) структура белковой молекулы [https://him.1september.ru/ 2004/06/12.htm].
Вторичной структурой называют пространственное расположение атомов главной цепи молекулы белка. Существует три типа вторичной структуры: альфа-спираль, бета-складчатость и бета-изгиб. Образуется и удерживается в пространстве за счет образования водородных связей между боковыми группировками АК основной цепи. Водородные связи образуются между электроотрицательными атомами кислорода карбонильных групп и атомами водорода двух аминокислот.
Альфа-спираль – это пептидная цепь штопорообразно закрученная вокруг воображаемого цилиндра. Диаметр такой спирали 0,5 А. В природных белках обнаружена только правая спираль. Некоторые белки (инсулин) имеют две параллельные спирали. Бета-складчатость – полипептидная цепь собрана в равнозначные складки. Бета-изгиб – образуется между тремя аминокислотами за счет водородной связи. Он необходим для изменения пространственного расположения полипептидной цепи при образовании третичной структуры белка.
Третичная структура – это свойственный данному белку способ укладки полипептидой цепи в пространстве. Это основа функциональности белка. Она обеспечивает стабильность обширных участков белка, состоящих из множества аминокислотных остатков и боковых групп. Такие упорядоченные в пространстве участки белка формируют активные центры ферментов или зоны связывания. Повреждение третичной структуры приводит к утрате функциональной активности белка.
Рис. 16.3. Третичная структура белковой молекулы, образованная за счет ковалентных связей между остатками двух молекул цистеина (дисульфидные мостики) [https://www.nsu.ru/education/biology/molbiol/Lecture3/Lec34.htm].
Стабильность третичной структуры зависит в основном от нековалентных взаимодействий внутри белковой глобулы – преимущественно водородных связей, ван-дер-ваальсовых сил (ионных или электростатических) и гидрофобных взаимодействий. Но некоторые белки дополнительно стабилизируются за счет таких ковалентных взаимодействий как дисульфидные мостики межу остатками цистеина.
Большинство белковых молекул имеют участки как альфа-спирали так и бета-складчатости. Но чаще по форме третичной структуры разделяют глобулярные белки – построенные преимуществено из альфа-спиралей и имеющеие форму шара или элипса (большинство ферментов). И фибрилярные – состоящие пеимущественно из бета-складчатости и имеющие сплющенную или нитевидную формы (пепсин, белки соединительной такни и хряща).
Размещение в пространстве взаимодействующих между собой субъединиц, образованных отдельными полипептидными цепями, называется четвертичной структурой. Т.е. в формировании четвертичной структуры участвуют не пептидные цепи сами по себе, а глобулы, образованные каждой из этих цепей в отдельности. Четвертичная структура – это высший уровень организации белковой молекулы и он присущ далеко не всем белкам. Связи, формирующие эту структуру нековалентные: водородные, электростатические.
Рис. 16.4. Четвертичная структура молекулы гемоглобина [https://www.nsu.ru/education/biology/molbiol/Lecture3/Lec35.htm].
Фундаментальный принцип молекулярной биологии: последовательность аминокислотных остатков полипептидной цепи белка несет в себе всю информацию, которая необходима для формирования определенной пространственной структуры. Т.е. имеющаяся в данном белке аминокислотная последовательность предопределяет образование альфа- или бета-конформации вторичной структуры за счет образования между этими АК водородных или дисульфидных связей и в дальнейшем формирование глобулярной или фибрилярной структуры также за счет нековалентных взаиомдействий между боковыми учатками определенных аминокислот.
Классификация белков
I. По растворимости:
§ водорастворимые,
§ солерстворимые,
§ спирторастворимые,
§ нерастворимые,
§ прочие.
II. По конформационной структуре:
§ фибриллярные (коллагены, эластины, кератины),
§ глобулярные: альбумины, глобулины, гистоны (все ферменты и большинство БАВ).
III. По химическому строению:
1. Протеины – состоят только из аминокислот.
a) Альбумины – растворимы в воде, не растворимы в концентрированных растворах солей.
b) Глобулины – не растворимы в воде, растворимы в солевых растворах.
c) Гистоны – растворимы в воде, в слабоконцентрированных кислотах. Обладают выраженными основными свойствами. Это ядерные белки, они связаны с ДНК и РНК.
d) Склеропротеины – белки опорных тканей (хрящей, костей), шерсти, волос. Не растворимы в воде, слабых кислотах и щелочах.
§ коллагены – фибрилярные белки соединительной ткани. При длительном кипячении они растворяются в воде и при застудневании образуется желатин.
§ эластины – белки связок и сухожилий. По свойствам похожи на коллагены, но подвергаются гидролизу под действием ферментов пищеварительного сока;
§ кератин – входит в состав волос;
2. Протеиды – помимо АК имеют в составе небелковую часть (глико-, липо-, металло-, фосфо-, нуклео-, хромопротеиды)
a) Нуклеопротеиды – простетическая група – нуклеиновые кислоты. Среди многочисленных классов нуклеопротеидов наиболее изученными являются рибосомы, состоящие из нескольких молекул РНК и рибосомных белков, и хроматин – основной нуклеопротеид эукариотических клеток, состоящий из ДНК и структурообразующих белков – гистонов (содержатся в клеточном ядре и митохондриях).
b) Гемопротеиды – небелковый компонент этих протеидов – гем, построен из четырех пиррольных колец, с ними связан ион двухвалентного железа (через атомы азота). К таким белка относятся: гемоглобин, миоглобин, цитохромы. Этот класс белков еще называют хромопротеиды, поскольку гем является окрашенным соединением. Гемоглобин – транспорт кислорода. Миоглобин – запасание кислорода в мышцах. Цитохромы (ферменты) – катализ окислительно-восстановаительных реакций и электронный транспорт в дыхательной цепи.
c) Металлопротеиды – в состав простетической группы входят металлы. Цитохром а – содержит медь, сукцинатдегидрогеназа и др. ферменты содержат негеминовое железо (ферродоксин).
d) Липопротеиды – содержат липиды, входят в состав клеточных мембран
e) Фосфопротеиды – содержат остаток фосфорной кислоты
f) Глюкопротеиды – содержат сахара