Гемоглобин его производные формы гемоглобина

Гемоглобин входит в состав группы белков гемопротеины, которые сами являются подвидом хромопротеинов и подразделяются на неферментативные белки (гемоглобин, миоглобин) и ферменты (цитохромы, каталаза, пероксидаза). Небелковой частью их является гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
Строение гемоглобина
Строение гемоглобина А
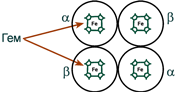
Гемоглобин представляет собой белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями, при этом они взаимодействуют не произвольно, а определенным участком – контактной поверхностью. Этот процесс высокоспецифичен, контакт происходит одновременно в десятках точек по принципу комплементарности. Взаимодействие осуществляют разноименно заряженные группы, гидрофобные участки, неровности на поверхности белка.
Белковые субъединицы в нормальном гемоглобине могут быть представлены различными типами полипептидных цепей: α, β, γ, δ, ε, ξ (соответственно, греч. – альфа, бета, гамма, дельта, эпсилон, кси). В состав молекулы гемоглобина входят по две цепи двух разных типов.
Гем соединяется с белковой субъединицей, во-первых, через остаток гистидина координационной связью железа, во-вторых, через гидрофобные связи пиррольных колец и гидрофобных аминокислот. Гем располагается как бы “в кармане” своей цепи и формируется гемсодержащий протомер.
Нормальные формы гемоглобина
Существует несколько нормальных вариантов гемоглобина:
- HbР (primitive) – примитивный гемоглобин, содержит 2ξ- и 2ε-цепи, встречается в эмбрионе между 7-12 неделями жизни,
- HbF (foetal) – фетальный гемоглобин, содержит 2α- и 2γ-цепи, появляется через 12 недель внутриутробного развития и является основным после 3 месяцев,
- HbA (adult) – гемоглобин взрослых, доля составляет 98%, содержит 2α- и 2β-цепи, у плода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина,
- HbA2 – гемоглобин взрослых, доля составляет 2%, содержит 2α- и 2δ-цепи,
- HbO2 – оксигемоглобин, образуется при связывании кислорода в легких, в легочных венах его 94-98% от всего количества гемоглобина,
- HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
Патологические формы гемоглобина
HbS – гемоглобин серповидно-клеточной анемии.
MetHb – метгемоглобин, форма гемоглобина, включающая трехвалентный ион железа вместо двухвалентного. Такая форма образуется спонтанно, при взаимодействии молекулы O2 и гемового Fe2+, но обычно ферментативных мощностей клетки хватает на его восстановление. При использовании сульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления Fe3+ в Fe2+ в клинике используют аскорбиновую кислоту и метиленовую синь.
Hb-CO – карбоксигемоглобин, образуется при наличии СО (угарный газ) во вдыхаемом воздухе. Он постоянно присутствует в крови в малых концентрациях, но его доля может колебаться от условий и образа жизни.
Угарный газ является активным ингибитором гем-содержащих ферментов, в частности, цитохромоксидазы, 4-го комплекса дыхательной цепи.
Карбоксигемоглобин присутствует и в норме в количестве 0,5-1,5%, в сельской местности меньше, чем в городе. У курильщиков концентрация Hb-CO возрастает, в зависимости от количества сигарет в день, до 8-9%.
HbA1С – гликозилированный гемоглобин. Концентрация его нарастает при хронической гипергликемии и является хорошим скрининговым показателем уровня глюкозы крови за длительный период времени (время жизни эритроцита, 3-4 месяца).
Функции гемоглобина
Строение гемоглобина
Синтез гемоглобина
Синтезированный в митохондриях гем индуцируется синтез цепей глобина на полирибосомах. Гены цепей глобина расположены в 11 и 16 хромосоме.
Цепи глобина формируют глобулы и соединяются с гемом. 4 глобулы нековалентно соединяются в гемоглобин.
Гемоглобин начинает синтезироваться на стадии базофильного эритробласта, а заканчивается у ретикулоцитов. В ретикулоцитах также идет синтез пуринов, пиримидинов, фосфатидов, липида. Чувствительным биохимическим индикатором для отличия ретикулоцитов от зрелых клеток является утрата последними глутаминазы. Глутамин в ретикулоцитах – источник углерода для синтеза порфирина и азота для синтеза пурина.
Гемоглобин – тетрамерный хромопротеин, имеет массу 64500Да, состоит из 4 гемов и 4 глобинов. Глобины представлены полипептидными цепями различных типов a, b, g, d и т.д. a-цепь содержит 141 АК, а b – цепь – 146 АК. Отдельные участки полипептидных цепей образуют правозакрученные a-спирали, особое расположение в пространстве которых формирует глобулы. Глобулаb-субъединицы содержит 8 a-спиралей, а a-субъединицы –7. Гем располагается в щелях между Е и F спиралями глобина, прикрепляясь через гистидин F8 к спирали F с помощью 5 координационной связи железа. Гидрофобные остатки аминокислот окружающие гем, препятствуют окислению железа водой. 4 глобулы с участием гидрофобных, ионных и водородных связей формируют шарообразный тетрамер гемоглобина. Максимально прочные связи, в основном за счет гидрофобных связей, образуются между a- и b-глобулами. В результате образуются 2 димера a1b1 и a2b2. Димеры соединяются между собой в основном полярными (ионными и водородными) связями, поэтому взаимодействие димеров зависит от рН. Димеры легко перемещаются друг относительно друга. В центре тетрамера глобулы прилегают друг к другу неплотно, образуя полость.
· Обеспечивают перенос кислорода от легких к тканям. В сутки около 600 литров;
· Участвует в переносе углекислого газа и протонов от тканей к легким;
· Регулирует КОС крови.
Гемоглобин со свободной шестой координационной связью железа в составе гема называется апогемоглобином.
Шестая координационная связь может связывать различные лиганды, с образованием следующих производных гемоглобина:
1) оксигемоглобин HbО2 (Fe2+) – соединение молекулярного кислорода с гемоглобином. Процесс называется оксигенацией; обратный процесс – дезоксигенацией.
2) карбоксигемоглобин HbСО (Fe2+). Связь гема с СО в двести раз прочнее, чем с О2. В норме в крови содержится 1% HbСО. У курильщиков к вечеру концентрация HbСО достигает 20%. При отравлении СО, из-за недостаточного снабжения тканей кислородом может наступить смерть.
3) метгемоглобин HbОН (Fe3+). Образуется при воздействии на гемоглобин окислителей (оксидов азота, метиленового синего, хлоратов). В норме в крови содержится <1% HbОН. Накопление метгемоглобина при некоторых заболеваниях (например, нарушение синтеза ГЛ-6-фосфатДГ), отравлении окислителями может стать причиной смерти, так как метгемоглобин не способен к переносу кислорода;
4) цианметгемоглобин HbСN (Fe3+). Образуется при присоединении СN- к метгемоглобину. Эта реакция спасает организм от смертельного действия цианидов. Поэтому для лечения отравлений цианидами применяют метгемоглобинообразователи (нитрит Na);
Карбгемоглобин образуется, когда гемоглобин связывается с СО2. Однако СО2 присоединяется не к гему, а к NН2 – группам глобина, с образованием карбаматов:
HbNH2 + CO2= HbNHCOO- + H+
Карбгемоглобин выводит из организма 10-15% СО2.
Дезоксигемоглобин Hb (Fe2+). Форма гемоглобина не связанная с кислородом. Дезоксигемоглобин связывает больше СО2, чем оксигемоглобин.
В цитохромах гем присоединяется к белковой части через 5 и 6 координационные связи железа (через гистидин и метионин Е и F спиралей). Занятость всех координационных связей не позволяет цитохромам присоединять лиганды, поэтому они могут переносить только по 1 электрону.
Гемоглобин со свободной шестой
координационной связью железа в составе
гема называется апогемоглобином.
Шестая координационная связь может
связывать различные лиганды, с образованием
следующих производных гемоглобина:
оксигемоглобинHbО2(Fe2+) – соединение
молекулярного кислорода с гемоглобином.
Процесс называется оксигенацией;
обратный процесс – дезоксигенацией.карбоксигемоглобинHbСО
(Fe2+). Связь гема с
СО в двести раз прочнее, чем с О2.
В норме в крови содержится 1%HbСО.
У курильщиков к вечеру концентрацияHbСО достигает 20%. При
отравлении СО, из-за недостаточного
снабжения тканей кислородом может
наступить смерть.метгемоглобинHbОН
(Fe3+). Образуется
при воздействии на гемоглобин окислителей
(оксидов азота, метиленового синего,
хлоратов). В норме в крови содержится<1%HbОН.
Накопление метгемоглобина при некоторых
заболеваниях (например, нарушение
синтеза ГЛ-6-фосфатДГ), отравлении
окислителями может стать причиной
смерти, так как метгемоглобин не способен
к переносу кислорода;цианметгемоглобинHbСN(Fe3+). Образуется
при присоединении СN-к метгемоглобину. Эта реакция спасает
организм от смертельного действия
цианидов. Поэтому для лечения отравлений
цианидами применяют метгемоглобинообразователи
(нитритNa);
Карбгемоглобинобразуется,
когда гемоглобин связывается с СО2.
Однако СО2присоединяется не к
гему, а кNН2– группам
глобина, с образованием карбаматов:
HbNH2 +CO2=HbNHCOO-+H+
Карбгемоглобин выводит из организма
10-15% СО2.
ДезоксигемоглобинHb(Fe2+). Форма гемоглобина
не связанная скислородом.
Дезоксигемоглобин связывает больше
СО2, чем оксигемоглобин.
В цитохромахгем присоединяется
к белковой части через 5 и 6 координационные
связи железа (через гистидин и метионин
Е иFспиралей). Занятость
всех координационных связей не позволяет
цитохромам присоединять лиганды, поэтому
они могут переносить только по 1 электрону.
Механизм насыщения гемоглобина
кислородом
Гемоглобин присоединяет О2последовательно, по одной молекуле на
каждый гем.
В апогемоглобине,благодаря
координационной связи с белковой частью,
атом железа выступает из плоскости гема
в направлении гистидинаF8.
Присоединение О2к шестой
координационной связи железа вызывает
его перемещение в плоскость гема, за
ним перемещаются гистидинF8
и полипептидная цепь, в состав которой
он входит.
Происходит изменение конформации
текущего протомера и связанных с ним
оставшихся протомеров. При этом у
протомеров возрастает сродство к
кислороду, в результате каждый следующий
кислород присоединяется к гемоглобину
лучше предыдущего. Четвертая молекула
кислорода присоединяется к гемоглобину
в 300 раз легче, чем первая молекула.
Обратный процесс аналогичен, чем больше
О2отдают протомеры, тем легче
идет отщепление последующих молекул
О2.
Кривая диссоциации кислорода для
гемоглобина
Кооперативность в работе протомеров
гемоглобина формирует сигмовидный
характер кривой насыщения его кислородом
в зависимости от парциального давления
кислорода.
S–образная кривая насыщения
гемоглобина кислородом имеет важное
биологическое значение.
Во-первых, пологий участокS–образной
кривой (выше 60мм.рт.ст.)
обеспечивает максимальное насыщение
гемоглобина кислородом в легких, даже
если концентрация кислорода в альвеолярном
воздухе заметно снижена. Например, в
альвеолярной крови приРО2=95
мм.рт.ст. гемоглобин насыщается кислородом
на 97%, а при РО2=60
мм.рт.ст. – на 90%.
Во-вторых,Крутой наклон
среднего участка S–образной
кривой (от 10 до40 мм.рт.ст.)
обеспечивает максимальный переход
кислорода от гемоглобина к тканям.
В области венозного конца
капилляра приРО2
= 40 мм.рт.ст. гемоглобин насыщен кислородом
на 73%. При снижении РО2
на 5 мм.рт.ст. насыщение гемоглобина
кислородом уменьшается на 7%.
Аллостерическая регуляция насыщения
гемоглобина кислородом
Кроме РО2
на насыщение гемоглобина
кислородом влияют и другие факторы,
например, рН, температура, давление,
концентрация 2,3-ДФГ,РСО2.
Увеличение температуры, присоединение
к гемоглобину Н+, 2,3-ДФГ, СО2уменьшает сродство гемоглобина к
кислороду, при этом кривая диссоциации
оксигемоглобина сдвигается вправо и
гемоглобин легче отдает кислород тканям.
Эффект Бора
Влияние рН на характер кривой диссоциации
оксигемоглобина называется эффектом
Бора(по имени датского физиолога
Христиана Бора, впервые открывшего этот
эффект).
Гемоглобин в дезоксигенерированном
состоянии имеет более высокое сродство
к протонам, чем оксигемоглобин. Другими
словами R – форма (оксигенерированная)
является более сильной кислотой, чем
Т-форма (дезоксигенерированная). Поэтому
когда дезоксигемоглобин в легких
присоединяет кислород, происходит
переход в R – форму и разрыв некоторых
связей, в результате чего и высвобождаются
протоны, ответственные за эффект Бора.
Наоборот, при высвобождении кислорода
образуется Т-структура и разорванные
связи между субъединицами должны быть
восстановлены, и протоны вновь
присоединяются к остаткам гистидина
в - цепях. Таким
образом, протонирование гемоглобина
снижает его сродство к О2 и увеличивает
потребление О2 в ткани.
Эффект Бора имеет важное физиологическое
значение. Образующийся в тканях СО2
должен транспортироваться в легкие. Он
поступает в эритроциты по градиенту
напряжения. В них фермент карбоангидраза
превращает его в Н2СО3, который диссоциирует
на бикарбонат, ион и протон. Последний
сдвигает равновесие влево в уравнении
(1).
Hb + 4 O2= Hb (О2)4 + (H+)n
Где n – величина порядка 2; число зависит
от целого комплекса параметров, тем
самым заставляя Hb О2 отдавать свой
кислород.
НСО3- пассивно продвигается через ионный
канал по градиенту концентрации в
сыворотку.
Продвижение НСО3- не сопровождается
перемещением Н+, поскольку нет канала,
позволяющего ему пройти через мембрану
эритроцитов. Для сохранения ионного
равновесия при выходе НСО3- из клетки,
Cl- перемещаются внутрь её через тот же
ионный канал. Такое двойное перемещение
известно как хлоридный сдвиг (сдвиг
Хамбургера).
Растворенный НСО3- движется вместе с
венозной кровью обратно в легкие. Здесь
высвобождение протона из гемоглобина
при оксигениции приводит к образованию
НСО3- (по принципу Ле-Шателье).
НСО3-+ Н+= Н2СО3-,
что позволяет карбоангидразе образовать
СО2.
Разрушение НСО3- в эритроците обуславливает
вхождение в него НСО3- из сыворотки, так
что в легких происходит обратный
хлоридный сдвиг, приводящий к выведению
СО2 с выдыхаемым воздухом.
Аллостерическая регуляция сродства
гемоглобина к кислороду 2,3-ДФГ
2,3-ДФГ снижает сродство гемоглобина к
кислороду и, таким образом, повышает
отдачу кислорода тканям. Если кровь
израсходовала весь свой запас ДФГ,
гемоглобин остается фактически насыщенным
кислородом. При акклиматизации в условиях
высокогорья содержание ДФГ в эритроцитах
резко увеличивается. ДФГ является
аллотерическим лигандом, так как
связывается с гемоглобином в другом по
сравнению с О2 участком. ДФГ встраивается
в полость тетрамерной молекулы
гемоглобина, полость образована остатками
всех 4 протомеров.
В Т – форме (дезоксигенерированной)
молекулы Hb имеются дополнительные
связи, и поэтому размер центральной
полости больше, чем в R – форме
(дезоксигемоглобине). Поэтому ДФГ
взаимодействует только с Т – формой
стабилизируя её, путем образования
связи между атомами кислорода ДФГ и
тремя положительно заряженными группами
в каждой из - цепей.
В легких при высоком парциальном давлении
кислород взаимодействует с Hb, изменяется
конформация белка, уменьшается центральная
полость и ДФГ вытесняется из гемоглобина.
Виды гемоглобинов
Гемоглобины различаются по белковой
части. Бывают физиологические и аномальные
виды гемоглобинов. Физиологические
образуются на разных этапах нормального
развития организма, а аномальные –
вследствие нарушения последовательности
аминокислот в глобине физиологических
видов гемоглобина.
Физиологические виды гемоглобина
1) эмбриональные гемоглобины(Gover I, Gover II). На ранних этапах развития
плода в первые недели развития, когда
в желточном мешке возникают очаги
кроветворения идет синтез гемоглобина
Gover I (состоит из четырёх эпсилон цепей:
4). Затем у эмбриона,
длина которого не превышает 2,5 см,
начинается синтез-цепей,
образуется гемоглобин Gover II (22). Эти гемоглобины
полностью исчезают у трехмесячного
эмбриона. Если они остаются у новорожденного,
то это признак врожденной аномалии
развития.
2) фетальный гемоглобин– HbF (от
латинского fetus – плод). Фетальный
гемоглобин сменяет эмбриональные
гемоглобины, вместо эпсилон – цепей (- цепей) начинают синтезироваться
гамма-цепи (- цепи).
HbF состоит из 2и 2цепей. HbF – является главным гемоглобином
плода и составляет к моменту рождения
50-80% всего гемоглобина. HbF имеет более
высокое сродство к кислороду, что
позволяет ему забирать кислород от
гемоглобина матери и передавать его
тканям плода. Эта особенность связана
с низким сродством HbF к 2,3-ФГК.
Кроме перечисленных основных видов
гемоглобинов плода, у здорового плода
выделяются и другие виды гемоглобинов:
например, гемоглобин Bart`s, (4),
Portland–1 (S22).
Схема
электрофореза
гемоглобина
здорового плода
+
А1
F
А2
Gower I







Bart`s
Portland -1
Gower II
3) гемоглобин А1–
тетрамер (22)
составляет около 98% гемоглобина
эритроцитов взрослого человека. Начинает
синтезироваться на 8 месяце развития
плода.
4) гемоглобин А2–
тетрамер (22).
Его содержание в эритроцитах взрослого
человека равно 2%. Гемоглобин А2,
также как и гемоглобин F, обладает более
высоким сродством к кислороду по
сравнению с гемоглобином А1.
5) гемоглобин А3(22)
образуется по мере старения эритроцита,
при присоединении к цистеину-цепи
глутатиона.
6) гемоглобин А1С– гликозилированный гемоглобин А.
Аномальные виды гемоглобинов
Аномальные гемоглобины возникают в
результате мутации генов, кодирующих
ицепи. Известно несколько сотен мутантных
гемоглобинов человека (в большинстве
случаев функционально активных).
Таблица №1 замена аминокислот в ипептидных цепях
гемоглобина
тип гемоглобина | нормальный | замена |
С | глу 6 в - цепи | лиз |
Д | лей 28 в - цепи | глу |
Е | глу 26 в - цепи | лиз |
G | глу 43 в - цепи | ала |
GpH | асл 68 в - цепи | лиз |
J | лиз 16 в - цепи | асл |
М | вал 67 в - цепи | глу |
О | глу 116 в - цепи | лиз |
S | глу 6 в - цепи | вал |
Болезни гемоглобинов
Болезни гемоглобинов называют
гемоглобинозами, их насчитывают
более 200.
Гемоглобинозы делятся на гемоглобинопатии
и таласемии.
Гемоглобинопатии, возникают в
результате точечных мутаций в структурных
генах, кодирующих полипептидные цепи
гемоглобина. Поэтому в крови появляется
аномальный гемоглобин.
Серповидноклеточная анемия–
классический пример наследственной
гемоглобинопатии. В норме в-субъединицах
гемоглобина в шестом положении находится
гидрофильная глутаминовая кислота. В
гемоглобине S глутаминовая кислота
заменена на гидрофобный валин. Такая
замена приводит к появлению на поверхности-субъединицы
гидрофобного («липкого») участка, который
соединяется с гидрофобным карманом
другой молекулы гемоглобина S. Происходит
полимеризация гемоглобина S и его
осаждение в виде длинных волокон. Длинная
волокнистая структура нарушает нормальную
форму эритроцитов, превращая её из
двояковогнутого диска в серповидную,
которая имеет тенденцию блокировать
капилляры. Такие эритроциты преждевременно
разрушаются, способствуя развитию
анемии. Если поражены обе гомологичные
хромосомы, заболевание может оказаться
смертельным. Заболевание широко
распространено в географических зонах,
где наиболее часто встречается
злокачественная форма малярии. Высокий
показатель заболеваемости можно
объяснить положительной селекцией
генома носителей аномальных генов.
Серповидная красная кровяная клетка
«неудобна» для развития малярийного
плазмодия.
Существенное ухудшение состояния
больных наблюдается в условиях высокогорья
при низких давлениях кислорода. Это
связано с тем, что полимеризоваться
способна только дезоксиформа S гемоглобина.
Так как в молекуле оксиформы S-гемоглобина
нет гидрофобного кармана («липкого
участка»), и она не способна к полимеризации.
Талассемия– генетическое заболевание,
обусловленное отсутствием или снижением
синтеза одной из цепей гемоглобина. При
данном заболевании отсутствуют дефекты
в структурных генах, кодирующих,,,-цепи.
Причиной талассемий являются мутации
генов-операторов, контролирующих
транскрипцию структурных генов ,,,-цепей гемоглобина.
В результате несбалансированного
образования глобиновых цепей образуются
тетрамеры гемоглобина, состоящие из
одинаковых протомеров.
В зависимости от того, формирование
какой глобиновой цепи нарушается,
выделяют ,
, ,
–
талассемии.
Талассемии делятся так же на гомозиготныеигетерозиготные.
Гомозиготная -талассемия– формирование-цепи
полностью подавляется. Симптомы
заболевания появляются приблизительно
через полгода после рождения, когда
происходит полное переключение синтеза-цепи гемоглобина
F на-цепь. У ребенка
прогрессирует анемия. Увеличиваются
селезенка и печень. Лицо приобретает
монголоидные черты (из-за чрезмерного
разрастания костного мозга скулы
выдаются вперед, нос приплюснут), при
рентгенологическом исследовании черепа
наблюдается феномен «игл ежа» («hair –
standing –on –end»). В попытке восполнить
эритроциты, утраченные в результате не
эффективного эритропоэза и увеличении
гемолиза, ткани черепа, чрезмерно
разрастаясь и гипертрофируясь, порождают
такое изменение медуллярной пластинки.
α-талассемия
– недостаток образования
α-глобиновых цепей приводит к нарушению
образования HbF
у плода. Избыточные
γ-цепи образуют тетрамеры, называемые
гемоглобином
Барта. Этот гемоглобин при физиологических
условиях имеет повышенное сродство
к кислороду и не проявляет кооперативных
взаимодействий между протомерами. В
результате гемоглобин Барта не
обеспечивает
развивающийся плод необходимым
количеством
кислорода, что приводит к тяжёлой
гипоксии. При α-талассемии
отмечают высокий процент внутриутробной
гибели плода.
Выжившие новорождённые при переключении
с γ- на β-ген синтезируют β-тетрамеры
или
НbН,
который, подобно гемоглобину Барта,
имеет слишком высокое сродство к
кислороду,
менее стабилен, чем НbА
и быстро разрушается.
Это ведёт к развитию у больных тканевой
гипоксии и к смерти вскоре после рождения.
Для всех этих заболеваний характерны
некоторые общие закономерности:
1). нарушаются пропорции в составе
гемоглобина крови. Например, при -
талассемии в крови появляется 15%
гемоглобина А2, 15 – 60% гемоглобина
F;
2). эритроциты приобретают не нормальную
форму (мишеневидную, каплевидную). Такие
эритроциты в пределах 1 дня захватываются
ретикулярной соединительной тканью
(например, селезенкой) и подвергаются
распаду (по этой причине селезёнка
оказывается гипертрофированной), что
приводит к развитию гемолитической
анемии.
Катаболизм гемоглобина
Старые поврежденные эритроциты
фагоцитируются клетками РЭС и
перевариваются в лизосомах. При распаде
гемоглобина образуется жёлчный пигмент
билирубин. Дальнейший катаболизм
билирубина в печени, кишечнике и почках
приводит к образованию уробилиногенов
и уробилина, которые выходятся с калом
и мочой. Железо, освобождающееся при
распаде гема, снова используется для
синтеза железосодержащих белков.
ПРАКТИЧЕСКАЯ ЧАСТЬ:
1. Количественное определение гемоглобина.
9. ЭТАЛОНЫ ОТВЕТОВ НА ЗАДАЧИ, тестовые
вопросы, используемые при контроле
знаний на занятии (можно в виде приложения)
10. ХАРАКТЕР И ОБЪЕМ ВОЗМОЖНОЙ
УЧЕБНО-ИССЛЕДОВАТЕЛЬСКОЙ РАБОТЫ ПО
ТЕМЕ
(Указать конкретно характер и форму
УИРС: подготовка реферативных выступлений,
проведение самостоятельных исследований,
имитационная игра, оформление истории
болезни с использованием монографической
литературы и др. формы)
11. ЛИТЕРАТУРА ДЛЯ ПОДГОТОВКИ К ЗАНЯТИЮ
ПРЕПОДАВАТЕЛЯМ: