Присоединение о2 к гемоглобину

Кооперативное взаимодействие
Взаимовлияние протомеров олигомерного белка друг на друга называется кооперативное взаимодействие.
В легких такое взаимодействие субъединиц гемоглобина повышает его сродство к кислороду и ускоряет присоединение кислорода в 300 раз. В тканях идет обратный процесс, сродство снижается и ускорение отдачи кислорода также 300-кратное.
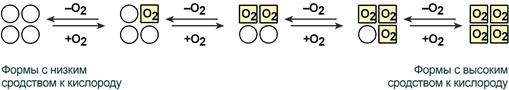
Схема кооперативного взаимодействия субъединиц гемоглобина
Объясняется такой феномен тем, что в легких при присоединении первой молекулы кислорода к железу (за счет 6-й координационной связи) атом железа втягивается в плоскость гема, кислород остается вне плоскости. Это вызывает перемещение участка белковой цепи и изменение конформации первого протомера. Такой измененный протомер влияет на другие субъединицы и облегчает связывание кислорода со второй субъединицей. Это меняет конформацию второй субъединицы, облегчая присоединение последующих молекул кислорода и изменение других протомеров.

Изменение формы субъединиц гемоглобина
при присоединении и отщеплении кислорода
Дезоксиформа гемоглобина обозначается как Т-форма, напряженная (англ. tense), она обладает существенно более низким сродством к кислороду. Оксигенированная форма, или R-форма (англ. relaxed), обладает высоким сродством к кислороду.
Изменение рН среды
Влияние рН на сродство гемоглобина к кислороду носит название эффекта Бора. При закислении среды сродство снижается, при защелачивании – повышается.
При повышении концентрации протонов (закисление среды) в тканях возрастает освобождение кислорода из оксигемоглобина. В легких после удаления угольной кислоты (в виде СО2) из крови и одновременном увеличении концентрации кислорода высвобождаются ионы Н+ из гемоглобина.
Реакция взаимодействия кислорода с гемоглобином упрощенно имеет вид:

Изменение сродства гемоглобина к кислороду в тканях и в легких при изменении концентрации ионов H+ и О2 обусловлено конформационными перестройками глобиновой части молекулы. В тканях молекула О2 отрывается от железа и ионы водорода присоединяются к остаткам гистидина (глобиновой части), образуя восстановленный гемоглобин (H-Hb) с низким сродством к кислороду. В легких поступающий в больших количествах кислород “вытесняет” ион водорода из связи с остатком гистидина гемоглобиновой молекулы.

Механизм эффекта Бора
Роль 2,3-дифосфоглицерата
2,3-Дифосфоглицерат образуется в эритроцитах из 1,3-дифосфоглицерата, промежуточного метаболита гликолиза, в реакциях, получивших название шунт Раппопорта.
Реакции шунта Раппопорта
2,3-Дифосфоглицерат располагается в центральной полости тетрамера дезоксигемоглобина и связывается с β-цепями, образуя поперечный солевой мостик между атомами кислорода 2,3-дифосфоглицерата и аминогруппами концевого валина обеих β-цепей, также аминогруппами радикалов лизина и гистидина.

Расположение 2,3-дифосфоглицерата в гемоглобине
Функция 2,3-дифосфоглицерата заключается в снижении сродства гемоглобина к кислороду, что имеет особенное значение при подъеме на высоту и при нехватке кислорода во вдыхаемом воздухе. В этих условиях связывание кислорода с гемоглобином в легких не нарушается, так как концентрация его относительно высока. Однако в тканях за счет 2,3-дифосфоглицерата отдача кислорода возрастает в 2 раза.
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 10 июля 2013;
проверки требуют 29 правок.
Эффект Вериго — Бора (синонимы — эффект Вериго, эффект Бора) — зависимость степени диссоциации оксигемоглобина от величины парциального давления углекислоты в альвеолярном воздухе и крови, при снижении которого сродство кислорода к гемоглобину повышается, что затрудняет переход кислорода из капилляров в ткани. Эффект этот был открыт независимо друг от друга Б. Ф. Вериго в 1892 году[1] и датским физиологом К. Бором в 1904 году[2].
История открытия[править | править код]
Основу разработки проблемы гипоксии заложил русский учёный-физиолог И. М. Сеченов фундаментальными работами по физиологии дыхания и газообменной функции крови. Большое значение имеют также исследования русского физиолога Б. Ф. Вериго по физиологии газообмена в лёгких и тканях. Опираясь на идеи Сеченова о сложных формах взаимодействия между диоксидом углерода и кислородом в крови (Вериго работал в лабораториях Сеченова, И. Р. Тарханова и И. И. Мечникова), он впервые установил зависимость степени диссоциации оксигемоглобина от величины парциального давления углекислоты в крови.
Биохимический механизм[править | править код]
Эффект Вериго — Бора: влияние pH на кривую насыщения гемоглобина кислородом
В клетках периферических тканей органическое топливо окисляется в митохондриях с использованием кислорода (клеточное дыхание), доставляемого гемоглобином из лёгких; при этом в качестве продуктов образуются углекислый газ, вода и другие соединения. Образование углекислого газа в тканях одновременно приводит и к повышению концентрации ионов H+ (то есть к понижению pH), поскольку при гидратации CO2 образуется H2CO3 — слабая угольная кислота, диссоциирующая на ионы H+ и бикарбонат-ионы:
H2CO3 ⇄ H+ + HCO3-.
Гемоглобин переносит значительную долю (около 20 %) общего количества CO2 и ионов H+, образующихся в тканях и поступающих в лёгкие и почки, обеспечивающих выделение этих продуктов.
За много лет до открытия этого механизма было обнаружено, что на связывание кислорода гемоглобином очень сильное влияние оказывает pH и концентрация CO2: при присоединении CO2 и ионов H+ способность гемоглобина связывать O2 снижается. Действительно, в периферических тканях с относительно низким значением pH и высокой концентрацией CO2 сродство гемоглобина к кислороду падает. И наоборот, в лёгочных капиллярах выделение CO2 и сопутствующее ему повышение pH крови приводит к увеличению сродства гемоглобина к кислороду. Это влияние величины pH и концентрации CO2 на связывание и освобождение O2 гемоглобином и называют эффектом Вериго — Бора.
Реакция связывания кислорода гемоглобином в виде
Hb + O2 ⇄ HbO2
в действительности отражает неполную картину, поскольку не учитывает дополнительные лиганды H+ и CO2.
Чтобы объяснить влияние концентрации ионов H+ на связывание кислорода, следует записать эту реакцию в иной форме:
HHb+ + O2 ⇄ HbO2 + H+,
где HHb+ — протонированная форма гемоглобина. Из этого уравнения следует, что кривая насыщения гемоглобина кислородом зависит от концентрации ионов H+. Гемоглобин связывает и O2, и ионы H+, но между этими двумя процессами существует обратная зависимость. Если парциальное давление кислорода велико (что наблюдается, например, в лёгких), то гемоглобин связывает его, освобождая при этом ионы H+. При низком парциальном давлении кислорода (что имеет место в тканях) связываться с гемоглобином будут ионы H+.
Эффективность связывания гемоглобином углекислого газа (с образованием карбаминогемоглобина, чаще именуемого карбгемоглобином[3]) находится в обратной зависимости от связывания кислорода. В тканях часть избыточного CO2 связывается с гемоглобином, сродство последнего к O2 снижается, и происходит высвобождение кислорода. В лёгких же связывается избыток O2 воздуха, тем самым сродство гемоглобина к CO2 уменьшается, и CO2 выделяется в альвеолярный воздух, способствуя лёгкому закислению крови за счёт ионов H+, возникающих при диссоциации угольной кислоты (см. выше).
Описанные зависимости делают молекулу гемоглобина великолепно приспособленной к осуществлению совместного переноса эритроцитами кислорода, углекислого газа и ионов H+[4].
См. также[править | править код]
- Эффект Холдейна (открыт Джоном Скоттом Холдейном)
Примечания[править | править код]
- ↑ Verigo B. F. Zur Frage über die Wirkung des Sauerstoff auf die Kohlensäureausscheidung in den Lungen // Archiv für die gesammte Physiologie des Menschen und der Thiere : статья. — 1892. — № 51. — С. 321—361.
- ↑ Chr. Bohr, K. Hasselbalch, and August Krogh. Concerning a Biologically Important Relationship — The Influence of the Carbon Dioxide Content of Blood on its Oxygen Binding // Skand. Arch. Physiol. : статья. — 1904. — № 16. — С. 401—412.
- ↑ Карбгемоглобин // Биологический энциклопедический словарь (рус.).
- ↑ Ленинджер А. Основы биохимии: В 3-х томах. Том 1. — М.: Мир, 1985. — С. 208—210. — 367 с.
Литература[править | править код]
- Вериго эффект // Большой медицинский словарь (рус.). — 2000.
Гемоглобин связывает и передает тканям за сутки около 600 л О2, в тканях образуется примерно 500 л СО2, который выводится из организма при участии гемоглобина.
Скорость оксигенирования и транспорт кислорода из лёгких в ткани зависит от изменений сродства гемоглобина к О2.
Присоединением О2 к одному из протомеров гемоглобина в лёгких при высоком рО2, (100 мм рт. ст.) вызывает конформационные изменения во всей олигомерной структуре гемоглобина и облегчает присоединение последующих молекул О2. Это явление носит название кооперативных изменений гемоглобина или кооперативного связывания О2, благодаря которому гемоглобин максимально связывает О2 в легких и легко отдает О2 в периферических тканях при более низких значениях рО2 (рис кривая насыщения миоглобина и гемоглобина).
Давление в капиллярах Давление в лёгочных
работающей мышцы а л ь в е о л а х
Рис.Кривые насыщения (диссоциации) миоглобина и гемоглобина кислородом.
Кривая насыщения для гемоглобина имеет сигмоидную форму (S-образную). Это указывает на кооперативное связывание О2 (в легких при высоком рО2) и освобождении О2 (в тканевых капиллярах при более низком рО2).
Эффект Бора
Гемоглобин не только переносит О2 от легких к периферическим тканям, но и ускоряет транспорт СО2 и протонов от тканей к легким.
Эффект Бора – это влияние концентрации СО2 и Н+ на сродство гемоглобина к О2.
Увеличение концентрации протонов и СО2 снижает сродство О2 к гемоглобину и усиливает транспорт О2 в ткани. Разные органы потребляют неодинаковые количества кислорода. Наиболее активно извлекают кислород из крови работающие мышцы (до 85%).
Если гемоглобин насыщен кислородом почти на 100%, то все молекулы гемоглобина находятся в R-форме, Нb(О2)4. Такое состояние гемоглобина возникает при высоком парциальном О2в капиллярах легких (100 мм рт. ст.) и при высоком сродстве Нb к О2. В капиллярах мышц и других тканей более низкое парциальное давление О2 , в этих условиях от гемоглобина могут отщепляться молекулы О2 и поступать в ткани. Например, при парциальном давлении кислорода в капиллярах 40 мм рт. ст. гемоглобин имеет насыщение ≈ 65%.
Далее действуют другие факторы, приводящие к снижению сродства гемоглобина к О2, тем самым увеличивая доставку кислорода в ткани:
а) гемоглобин протонируется, связывая три пары Н+ в пептидных цепях гемоглобина, вновь формируется Т-структура;
б) гемоглобин обратимо присоединяет часть СО2 (до 15%) к концевым аминогруппам пептидных цепей, образуется карбогемоглобин: R-NH-COO- +Н+, где R-белковая цепь гемоглобина. Это также снижает сродство гемоглобина к О2. Большая часть СО2 транспортируется кровью в виде бикарбоната НСО3 -.
Таким образом, диоксид углерода, образовавшийся в тканях реагирует с водой, образуя угольную кислоту, которая диссоциирует на бикарбонат- ион и Н+. Синтез углекислоты, как и её разложение на СО2 и Н2О происходит при участии фермента карбоангидразы. В периферических тканях гемоглобин отдает О2, сам протонируется и связывает часть СО2, его R-форма переходит в Т-форму, сродство к О2 резко снижается. В легких происходят обратные процессы.
Влияние 2,3-БФГ на сродство гемоглобина к кислороду
2,3-биcфосфоглицерат (2,3-дифосфоглицерат) – образуется в эритроцитах в ходе анаэробного гликолиза из промежуточного соединения 1,3-биcфосфоглицерата. 2,3-БФГ является аллостерическим регулятором гемоглобина (от греч. «аллос» – другой), он соединяется ионными связями с дезоксигемоглобином не в области гема, а в другом участке, т. е. аллостерическом центре, содержащим положительно заряженные радикалы аминокислот Лиз 82 и Гис 143 β-цепей.
Структура 2,3-бифосфоглицерата
Образовавшиеся дополнительные солевые мостики (или ионные связи) делают структуру дезоксигемоглобина ещё более жесткой и поэтому сродство Нb к О2 снижается. Концентрация 2,3-БФГ в крови близка к концентрации гемоглобина и одинакова в артериальной и венозной крови. 2,3-БФГ реагирует только с дезоксигемоглобином, т. е. Т-формой, т. к. он имеет доступную центральную полость с центром связывания 2,3-БФГ. В оксигемоглобине (R-форма) эта полость закрыта.
Оглавление темы “Вентиляция легких. Перфузия легких кровью.”: Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.Молекула гемоглобина может находиться в двух формах — напряженной и расслабленной. Расслабленная форма гемоглобина имеет свойство насыщаться кислородом в 70 раз быстрее, чем напряженная. Изменение фракций напряженной и расслабленной формы в общем количестве гемоглобина в крови обусловливает S-образный вид кривой диссоциации оксигемоглобина, а следовательно, так называемое сродство гемоглобина к кислороду. Если вероятность перехода от напряженной формы гемоглобина к расслабленной больше, то возрастает сродство гемоглобина к кислороду, и наоборот. Вероятность образования указанных фракций гемоглобина изменяется в большую или меньшую сторону под влиянием нескольких факторов. Основной фактор — это связывание кислорода с геминовой фуппой молекулы гемоглобина. При этом чем больше геминовых фупп гемоглобина связывают кислород в эритроцитах, тем более легким становится переход молекулы гемоглобина к расслабленной форме и тем выше их сродство к кислороду. Поэтому при низком Р02, что имеет место в метаболически активных тканях, сродство гемоглобина к кислороду ниже, а при высоком Р02 — выше. Как только гемоглобин захватывает кислород, повышается его сродство к кислороду и молекула гемоглобина становится насыщенной при связывании с четырьмя молекулами кислорода.
Когда эритроциты, содержащие гемоглобин, достигают тканей, то кислород из эритроцитов диффундирует в клетки. В мышцах он поступает в своеобразного депо кислорода — в молекулы миоглобина, из которого кислород используется в биологическом окислении мышц. Диффузия кислорода из гемоглобина эритроцитов в ткани обусловлена низким Р02 в тканях — 35 мм рт. ст. Внутри клеток тканей напряжение кислорода, необходимое для поддержания нормального метаболизма, составляет еще меньшую величину — не более 1 кПа. Поэтому кислород путем диффузии из капилляров достигает метаболически активных клеток. Некоторые ткани приспособлены к низкому содержанию Р02 в капиллярах крови, что компенсируется высокой плотностью капилляров на единицу объема тканей. Например, в скелетной и сердечной мышцах Р02 в капиллярах может снизиться чрезвычайно быстро во время сокращения. В мышечных клетках содержится белок миоглобин, который имеет более высокое сродство к кислороду, чем гемоглобин. Миоглобин интенсивно насыщается кислородом и способствует его диффузии из крови в скелетную и сердечную мышцы, где он обусловливает процессы биологического окисления. Эти ткани способны экстрагировать до 70 % кислорода из крови, проходящей через них, что обусловлено снижением сродства гемоглобина к кислороду под влиянием температуры тканей и рН. Эффект рН и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое рН понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и С02. Если рН плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном рН 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда рН снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 1/з больше, чем при постоянном рН.
Метаболически активные ткани повышают продукцию тепла. Повышение температуры тканей при физической работе изменяет соотношение фракций гемоглобина в эритроцитах и вызывает смещение кривой диссоциации оксигемоглобина вправо вдоль оси х. В результате большее количество кислорода будет освобождаться из гемоглобина эритроцитов и поступать в ткани. Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении Р02 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3-ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей. – Также рекомендуем “Углекислый газ. Транспорт углекислого газа.” |


Основная
функция гемоглобина – доставка О2 от
лёгких к тканям. Олигомерная структура
гемоглобина обеспечивает быстрое
насыщение его кислородом в лёгких
(образование оксигемоглобина – Нb(О2)4),
возможность отщепления кислорода от
гемоглобина в капиллярах тканей при
относительно высоком парциальном
давлении О2,
а также возможность регуляции сродства
гемоглобина к О2 в
зависимости от потребностей тканей в
кислороде.
Кооперативные
изменения конформации протомеров
О2 связывается
с протомерами гемоглобина через Fe2+ ,
который соединён с четырьмя атомами
азота пиррольных колец тема и атомом
азота
Рис.
1-32. Строение гемоглобина.
Гис
F8 белковой
части протомера. Связывание О2 с
оставшейся свободной координационной
связью Fe2+ происходит
по другую сторону от плоскости гема в
области Гис Е7 (аналогично
тому, как это происходит у миоглобина).
Гис Е7 не
взаимодействует с О2,
но обеспечивает оптимальные условия
для его связывания (рис. 1-33).
В
дезоксигемоглобине благодаря ковалентной
связи с белковой частью атом Fe2+ выступает
из плоскости гема в направлении Гис F8.
Присоединение О2 к
атому Fe2+ одного
протомера вызывает его перемещение в
плоскость гема, за ним перемещаются
остаток Гис F8 и
полипептидная цепь, в состав которой
он входит. Так как протомер связан с
остальными протомерами, а белки обладают
конформационной лабильностью, происходит
изменение конформации всего белка.
Конформационные изменения, произошедшие
в других протомерах, облегчают
присоединение следующей молекулы О2,
что вызывает новые конформационные
изменения в белке и ускорение связывания
следующей молекулы О2.
Четвёртая молекула О2 присоединяется
к гемоглобину в 300 раз легче, чем первая
молекула (рис. 1-34).
Рис.
1-33. Изменение прложения Fe2+ и
белковой части гемоглобина при
присоединении О2.
Рис.
1-34. Кооперативные изменения конформации
протомеров гемоглобина при присоединении
О2.
Изменение
конформации (а следовательно и
функциональных свойств) всех протомеров
олигомерного белка при присоединении
лиганда только к одному из них носит
название кооперативных изменений
конформации протомеров.
Аналогичным
образом в тканях диссоциация каждой
молекулы О2 изменяет
конформацию всех протомеров и облегчает
отщепление последующих молекул О2.
Кривые
диссоциации О2 для
миоглобина и гемоглобина
Кооперативность
в работе протомеров гемоглобина можно
наблюдать и на кривых диссоциации О2 для
миоглобина и гемоглобина (рис. 1-35).
Отношение
занятых О2 участков
связывания белка к общему числу таких
участков, способных к связыванию,
называется степенью насыщения этих
белков кислородом. Кривые диссоциации
показывают, насколько насыщены данные
белки О2 при
различных значениях парциального
давления кислорода.
Кривая
диссоциации О2 для
миоглобина имеет
вид простой гиперболы. Это указывает
на то, что миоглобин обратимо связывается
с лигандом, и на это не оказывают влияние
никакие посторонние факторы (схема
ниже).
Схема

Рис.
1-35. Кривые диссоциации кислорода для
миоглобина и гемоглобина в зависимости
от парциального давления кислорода.
Процессы
образования и распада оксимиоглобина
находятся в равновесии, и это равновесие
смещается влево или вправо в зависимости
от того, добавляется или удаляется
кислород из системы. Миоглобин связывает
кислород, который в капиллярах тканей
высвобождает гемоглобин, и сам миоглобин
может освобождать О2 в
ответ на возрастание потребностей в
нём мышечной ткани и при интенсивном
использовании О2 в
результате физической нагрузки.
Миоглобин
имеет очень высокое сродство к О2.
Даже при парциальном давлении О2,
равном 1-2 мм рт. ст., миоглобин остаётся
связанным с О2 на
50%.
Кривая
диссоциации О2 для
гемоглобина. Из
графика на рис. 1-35 видно, что гемоглобин
имеет значительно более низкое сродство
к О2;
полунасыщение гемоглобина О2 наступает
при более высоком давлении О2 (около
26 мм рт. ст.).
Кривая
диссоциации для гемоглобина имеет
сигмоидную форму (S-образную). Это
указывает на то, что протомеры гемоглобина
работают кооперативно: чем больше
О2 отдают
протомеры, тем легче идёт отщепление
последующих молекул О2.
В
капиллярах покоящихся мышц, где давление
О2 составляет
около 40 мм рт. ст., большая часть кислорода
возвращается в составе оксигемоглобина
обратно в лёгкие. При физической работе
давление О2 в
капиллярах мышц падает до 10-20 мм рт. ст.
Именно в этой области (от 10 до 40 мм рт.
ст.) располагается “крутая часть”
S-образной кривой, где в наибольшей
степени проявляется свойство кооперативной
работы протомеров.
Следовательно,
благодаря уникальной структуре каждый
из рассмотренных белков приспособлен
выполнять свою функцию: миоглобин –
присоединять О2,
высвобождаемый гемоглобином, накапливать
в клетке и отдавать в случае крайней
необходимости; гемоглобин – присоединять
О2 в
лёгких, где его насыщение доходит до
100%, и отдавать О2 в
капиллярах тканей в зависимости от
изменения в них давления О2
4.
Перенос Н+ и
С02 из
тканей в лёгкие
с помощью гемоглобина.
Эффект Бора
Окисление
органических веществ с целью получения
энергии происходит в митохондриях
клеток с использованием О2,
доставляемого гемоглобином из лёгких.
В результате окисления веществ образуются
конечные продукты распада – СО2 и
Н2О,
количество которых пропорционально
интенсивности процессов окисления.
СО2,
образовавшийся в тканях, транспортируется
в эритроциты. Там под действием фермента
карбангидразы происходит увеличение
скорости образования Н2СО3.
Слабая угольная кислота может
диссоциировать на Н+ и
НСО3-
СО2 +
Н2О
– H2CO 3 –
H+ +
HCO3-.
Равновесие
реакции в эритроцитах, находящихся в
капиллярах тканей, смещается вправо,
так как образующиеся в результате
диссоциации угольной кислоты протоны
могут присоединяться к специфическим
участкам молекулы гемоглобина: к
радикалам Гис146 двух
?-цепей, радикалам Гис122 и
концевым ?-аминогруппам двух ?-цепей.
Все эти 6 участков при переходе гемоглобина
от окси- к дезоксиформе приобретают
большее сродство к Н+ в
результате локального изменения
аминокислотного окружения вокруг этих
участков (приближения к ним отрицательно
заряженных карбоксильных групп
аминокислот).
Присоединение
3 пар протонов к гемоглобину уменьшает
его сродство к О2 и
усиливает транспорт О2 в
ткани, нуждающиеся в нём (рис. 1-36, А).
Увеличение освобождения О2 гемоглобином
в зависимости от концентрации Н+ называют
эффектом Бора (по имени датского физиолога
Христиана Бора, впервые открывшего этот
эффект).
В
капиллярах лёгких высокое парциальное
давление О2 приводит
к оксигенированию гемоглобина и удалению
6 протонов. Реакция СО2 +
Н2О
– Н2СО3 –
Н+ +
НСО3- сдвигается
влево и образующийся СО2 выделяется
в альвеолярное пространство и удаляется
с выдыхаемым воздухом (рис. 1-36, Б).
Следовательно,
молекула гемоглобина в ходе эволюции
приобрела способность воспринимать и
реагировать на информацию, получаемую
из окружающей среды. Увеличение
концентрации протонов в среде снижает
сродство О2 к
гемоглобину и усиливает его транспорт
в ткани (рис. 1-37).
Большая
часть СО2 транспортируется
кровью в виде бикарбоната НСО3-.
Небольшое количество
Рис.
1-36. Перенос Н+ и
СО2 с
кровью. Эффект Бора. А
– влияние концентрации СО2 и
Н+ на
высвобождение О2 из
комплекса с гемоглобином в тканях
(эффект Бора); Б – оксигенирование
дезоксигемоглобина в лёгких, образование
и выделение СО2.
Рис.
1-37. Влияние рН на кривую диссоциации
О2 для
гемоглобина.
СО2 (около
15-20%) может переноситься в лёгкие, обратимо
присоединяясь к неионизированным
концевым ?-аминогруппам. R-NH2+
СО2 =
R-NH-COO + Н+,
в результате образуется карбогемоглобин,
где R – полипептидная цепь гемоглобина.
Присоединение СО2 к
гемоглобину также снижает его сродство
к О2.
5. 2,3-Бифосфоглицерат
– аллостертеский регулятор сродства
гемоглобина к О2
2,3-Бифосфоглицерат
(БФГ) – вещество, синтезируемое в
эритроцитах из промежуточного продукта
окисления глюкозы 1,3-бифосфоглицерата.
2,3-Бифосфоглицерат
Регуляция
с помощью 2,3-бифосфоглицерата сродства
гемоглобина к О2
В
нормальньж условиях 2,3-бифосфоглицерат
присутствует в эритроцитах примерно в
той же концентрации, что и гемоглобин.
БФГ, присоединяясь к гемоглобину, также
может менять его сродство к О2.
В
центре тетрамерной молекулы гемоглобина
есть полость, образованная аминокислотными
остатками всех четырёх протомеров.
Центральная
полость – место присоединения БФГ.
Размеры
центральной полости могут меняться:
отщепление О2 от
оксигемоглобина вызывает его
конформационные изменения, которые
способствуют образованию дополнительных
ионных связей между димерами ?1?1 и
?2?2.
В результате пространственная структура
дезоксигемоглобина становится более
жёсткой, напряжённой, а центральная
полость расширяется.
Поверхность
полости ограничена остатками аминокислот,
в числе которых имеются положительно
заряженные радикалы Лиз82,
Гис143 ?-цепей
и положительно заряженные ?-аминогруппы
N-концевого валина ?-цепей. В расширенную
полость дезоксигемоглобина БФГ, имеющий
сильный отрицательный заряд, присоединяется
с помощью ионных связей, образующихся
с положительно заряженными функциональными
группами двух ?-цепей гемоглобина.
Присоединение БФГ ещё сильнее стабилизирует
жёсткую структуру дезоксигемоглобина
и снижает сродство белка к О2 (рис.
1-38).
Присоединение
БФГ к дезоксигемоглобину происходит в
участке, ином по сравнению с ге-мом, где
происходит связывание О2.
Такой лиганд называется “аллостерический”, а
центр, где связывается аллостерический
лиганд, –“аллостерический
центр” (от
греч. “аллос” – другой, иной, “стерос”
– пространственный).
В
лёгких высокое парциальное давление
О2 приводит к оксигенированию гемоглобина.
Разрыв
Рис.
1-38. Взаимодействие 2,3-бифосфоглицерата
с аминокислотными остатками центральной
полости дезоксигемоглобина.
ионных
связей между димерами ?1?1 и
?2?2 приводит
к “расслаблению” белковой молекулы,
уменьшению центральной полости и
вытеснению БФГ.
Изменение
концентрации БФГ как механизм адаптации
организма к гипоксии. Концентрация
БФГ в эритроцитах людей, живущих в
определённых климатических условиях,
– величина постоянная. Однако в период
адаптации к высокогорью, когда человек
поднимается на высоту более 4000 м над
уровнем моря, концентрация БФГ уже через
2 дня возрастает почти в 2 раза (от 4,5 до
7,0 мМ). Это снижает сродство гемоглобина
к О2 и
увеличивает количестве О2,
транспортируемого в ткани (рис. 1-39).
Такую
же адаптацию наблюдают у больных с
заболеваниями лёгких, при которых
развивается общая гипоксия тканей. Так,
у больных с тяжёлой обструктивной
эмфиземой лёгких парциальное давление
в них снижается от 100 до 50 мм рт. ст. Но
при этом в эритроцитах усиливается
выработка БФГ, и его концентрация
повышается с 4,5 до 7,0 мМ, что существенно
увеличивает доставку О2 в
ткани.
Рис.
1-39. Влияние различных концентраций
2,3-бифосфоглицерата на сродство
гемоглобина к О2.
Клиническое
значение концентрации БФГ в консервированной
крови
В
крови, консервированной в некоторых
средах, например цитратдекстрозной, за
10 дней концентрация БФГ снижается с 4,5
до 0,5 мМ. Гемоглобин такой крови имеет
очень высокое сродство к О2.
Если кровь со сниженной концентрацией
БФГ переливать тяжелобольным, возникает
опасность развития гипоксии тканей.
Введённые с кровью эритроциты за 24 ч
могут восстановить лишь половину
нормальной концентрации БФГ. Добавлением
в кровь БФГ нельзя восстановить нормальную
концентрацию его в эритроцитах, так
как, имея высокий отрицательный заряд,
БФГ не может проникать через мембраны
эритроцитов. Поэтому в настоящее время
в кровь добавляют вещества, способные
проникать через мембрану эритроцитов
и поддерживать в них нормальную
концентрацию БФГ.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #