Распад гемоглобина в организме человека

Происходит в мононуклеарных фагоцитах (в селезенке и печени). Сначала происходит разрыв связи между I-м и II-м кольцами. Фермент – НАДзависимая оксигеназа. После ее действия образуется пигмент зеленого цвета – вердоглобин. В его составе сохраняется и железо и белок. В дальнейшем вердоглобин распадается на составные части: белок, железо и пигмент-биливердин. Железо образует комплекс с белком трансферрином, и этот комплекс поступает в ткани, где оно снова может быть использовано для синтезов или депонироваться в виде комплекса с белком ферритином.
Билирубин плохо растворим в воде, поэтому он адсорбируется на альбумине крови. Такой комплекс называется СВОБОДНЫМ или НЕПРЯМЫМ БИЛИРУБИНОМ. Он свободный, потому что не связан ни с каким другим веществом ковалентными связями; непрямой, потому что не дает прямой реакции с диазореактивом Эрлиха (для определения этого вида билирубина необходимо предварительно осадить альбумины).
В печени непрямой билирубин подвергается обезвреживанию путем связывания с глюкуроновой кислотой. В реакции расходуется УТФ. При этом к молекуле билирубина присоединяются 2 молекулы глюкуроновой кислоты (образование билирубиндиглюкуронида). Это вещество малотоксично и хорошо растворяется в воде, называется ПРЯМЫМ СВЯЗАННЫМ БИЛИРУБИНОМ, т.к. определяется в прямой реакции с диазореактивом Эрлиха и связан ковалентно с глюкуроновой кислотой. В таком виде билирубин поступает в желчь и вместе с ней выводится из организма.
При определении содержания билирубина используют диазореактив Эрлиха, который дает розовую окраску с билирубином и по ее интенсивности судят о концентрации билирубина в крови.
В крови человека примерно 75% непрямого билирубина от его общего количества. Общее содержание билирубина в норме (прямой+непрямой) от 8 до 20 мкмоль/л.
Билирубин – это конечный продукт распада гемоглобина у человека. Однако, поступив с желчью в кишечник, он превращается под действием ферментов микрофлоры кишечника. Сначала от него отщепляется глюкуроновая кислота, затем происходит частичное восстановление и образуется вещество, называемое “мезобилиноген”, а затем стеркобилиноген. Стеркобилиноген в основном выводится с калом и на воздухе (в кале) превращается в стеркобилин, а мезобилиноген частично всасывается из кишечника по системе v.portae, т.е. попадает в печень и там разрушается. Поэтому его нет у здоровых людей в крови. Небольшая часть стеркобилиногена тоже всасывается из кишечника через систему геморроидальных вен, т.е. попадает сразу в большой круг кровообращения, минуя печень, поэтому часть его выводится с мочой.
Значит, стеркобилиноген является компонентом мочи в норме.
В НОРМЕ:
Кровь:
Билирубин (в основном непрямой) – в норме (8-17мкмоль/л)
Гемоглобин – в норме (мужчины – 130-145 г/л; женщины – 110-130 г/л
Моча: светло-желтого цвета, стеркобилиноген – в норме, мезобилиногена нет.
Кал: нормальной окраски, стеркобилиноген – в норме
135.
ГЕМОЛИТИЧЕСКАЯ ЖЕЛТУХА Наблюдается при усилении распада эритроцитов. Билирубина образуется больше, чем в норме и скорость экскреции тоже увеличивается. Непрямой билирубин повышается в крови. В моче билирубина нет, а содержание стеркобилина в кале и в моче повышено.
136.
ПАРЕНХИМАТОЗНАЯ ЖЕЛТУХА (паренхиматозная). Наблюдается при повреждении гепатоцитов (например, при вирусном гепатите). Билирубин не обезвреживается. Желчные пигменты поступают не только в кишечник, но и в кровь. Т.е. в крови повышается содержание не только прямого, но и непрямого билирубина. Наблюдается билирубинурия. В моче обнаруживают мезобилиноген.
137.
ОБТУРАЦИОННАЯ ЖЕЛТУХА (механическая) Наблюдается при закупорке желчных протоков (например, при желчнокаменной болезни). Желчь продолжает вырабатываться, но поступает не только в желчные протоки, а также в кровь. В крови повышается уровень билирубина, в основном – прямого билирубина. Наблюдается билирубинурия. Содержание стеркобилина в моче и в кале снижено (кал становится светлым, а моча темнеет за счет прямого билирубина).
138.
Лекция № 9.
Тема «ОБМЕН ХРОМОПРОТЕИДОВ».
План изложения:
1.Хромопротеины. Гемоглобин и миоглобин. Строение, функции, биологическое значение для организма, отличия.
2.Оксигенирование гемоглобина и миоглобина.
3.Синтез гема.
4.Распад гемоглобина в организме.
5.Клиническое значение исследования пигментов. Лабораторные показатели желтух.
К хромопротеинам относятся гемоглобин, миоглобин, хлорофилл.
Гемоглобин и миоглобин – сложные белки класса хромопротеинов подгруппы гемопротеинов, так как небелковой их частью является гемм.
Уникальным свойством этих белков является обратимое связывание с молекулами кислорода без изменения степени окисления иона железа
(Fe 2 +).
Отличаются эти белки локализацией и функциями.
1.Гемоглобин находится в эритроцитах, транспортирует кислород из лёгких к тканям, а из тканей в лёгкие переносит углекислый газ и протоны водорода. Эта его функция тесно связана с регуляцией кислотно-основного состояния в организме.
2.Миоглобин локализуется в цитоплазме мышечных клеток (красные мышцы), особенно много его в миокарде, сохраняет запас кислорода для анаэробного митохондриального окисления при интенсивной мышечной работе и транспортирует кислород внутриклеточно к митохондриям.
3.Сродство миоглобина к кислороду выше, чем у гемоглобина.
Различия в функциях этих белков обусловлены разной структурной организацией их молекул.
1.Миоглобин состоит из одной полипептидной цепи, содержащей 153 остатка аминокислот.
2.Гемоглобин имеет четыре попарно идентичные полипептидные цепи. У основного гемоглобина взрослых HbA комбинируются 2 альфа цепи, состоящие из 141 остатка аминокислот, и 2 вета-цепи, содержащие по 146 остатков.
3.Третичная конформация этих цепей очень похожа на аналогичную в молекуле миоглобина с той разницей, что в альфа цепи имеется семь спирализованных участков.
Каждая полипептидная цепь, соединённая с молекулой гема, образует субъединицу. Четыре субъединицы, взаимодействуя между собой нековалентными связями, укладываются в форме тетраэдра, образуя глобулярную четвертичную структуру.
Присоединение молекулярного кислорода к гемоглобину и миоглобину осуществляется через ион железа гема. Этот процесс называют оксигенированием(не окислением, так как он не приводит к изменению степени окисления железа!). Оксигенированные белки называют оксигемоглобинами (HbO 2) и оксимиоглобинами (MyoHbO 2).
На процесс оксигенирования гемоглобина влияют:
1.парциальное давление кислорода, углекислого газа,
2.концентрация протонов водорода (pH)
3.концентрация 2,3 – дифосфоглицерата (2,3- ДФГ) – промежуточного метаболита гликолиза в эритроцитах.
Четвертичная структура гемоглобина наделяет гемоглобин уникальной способностью насыщаться кислородом постепенно, достигая максимума при высоком парциальном давлении, и отдавать часть кислорода в ткани при снижении pO 2.
Пусковым механизмом изменения конформации гемоглобина при взаимодействии с О 2 при его высоком давлении служит перемещение иона железа в плоскость порфиринового кольца.
Максимальное насыщение кислородом ( на 96%) происходит в эритроцитах лёгочных капилляров при pO 2 = 100 мм рт.ст. В венозной крови гемоглобин насыщен кислородом только на 64%, следовательно 32% кіслорода переходит в ткани. Другими словами, на уровне периферических тканей происходит снижение сродства гемоглобина к кислороду и частичное дезоксигенирование.
Этому способствуют следующие условия:
1.низкое парциальное давление кислорода в периферических тканях (40 мм рт.ст.), а в работающих мышцах 20 мм рт. ст., так как в тканях кислород быстро используется на окислительное фосфорилирование для получения АТФ и некоторые другие процессы;
2.высокое pCO 2 образующееся в тканях в результате декарбоксилирования альфа- кетокислот, аминокислот, других карбоновых кислот в процессе метаболизма.
3. увеличение концентрации 2,3 – ДФГ, стимулируемое высоким рCO 2 Молекула оксігемоглобіна связывается определённым центром с молекулой 2, 3- дифосфоглицерата, что приводит к изменению конформации и освобождению части О 2.
Таким образом, оксигемоглобин снабжает ткани кислородом.
Оксигенирование миоглобина в мышцах и быстро и при низком парциальном давлении О 2 (20мм рт. ст.). Сродство миоглобина к кислороду выше, чем у гемоглобина. При интенсивной мышечной работе, приводящей к быстрому кислородному голоданию (рО 2 снижается до 5 мм рт. ст.) в клетке, оксимиоглобин переносит О 2, к митохондриям и отдаёт его в матрикс, где находится фермент цитохромоксидаза, имеющий более высокое сродство
к О 2, чем гемоглобин., и использующий кислород в качестве акцептора электронов.
СИНТЕЗ ГЕМА.
Гемоглобин синтезируется на стадии созревания эритроцитов. Он состоит из 4 полипептидов и 4-х молекул гема. Синтез гема происходит в митохондриях клеток костного мозга, печени, почках, слизистой кишечника, в ретикулоцитах. Субстратом дла синтеза гема являются глицин и сукцинил- КоА.
В6
Глицин +сукцинил – КоА порфобилиноген протопорфирин IX Fe 2+ гемм.
Транспорт гема в плазме крови осуществляется белком гемопексином. Это предотвращает повреждение почечного фильтра прооксидантом Fe 2+ и сохраняет гемм и Fe 2+ в организме.
Распад гемоглобина в организме.
Поступающий с пищей Hb в ЖКТ распадается на белковую часть – глобин и небелковую – гем. Глобин распадается до аминокислот, которые всасываются и поступают в кровь. Гем окисляется в гематин и выводится с калом, т е. не используется организмом.
Эндогенный гемоглобин распадается следующим образом:
процесс распада Er протекает в селезёнке, к/м, печени, СМФ (купферовские клетки, плазматические клетки, гистиоциты). Часть Er распадается в кровеносном русле. Освободившийся при этом Hb (около 8 – 9г), адсорбируется в крови гаптоглобином, одна молекула которого может связывать две молекулы гемоглобина. Образующийся комплекс адсорбируется клетками ретикулоэндотелиальной системы (РЭС) из крови, циркулирующей в печени, селезёнке, костном мозге и других органах.
Hb, окисляясь при помощи оксигеназы превращается в вердоглобин (пигмент зелёного цвета), вердоглобин нестойкое соединение, которое распадается на Fe (захватывается трансферрином и в печени откладывается в виде ферритина) и глобин ( гидролиз до аминокислот).
Образуется биливердин, который восстанавливается в эндоплазматическом ретикулуме клеток Купфера в присутствие биливердин – редуктазы в основной желчный пигмент человека -билирубин, так называемый «непрямой билирубин» (пигмент оранжевого цвета, токсичный, водонерастворимый).
«Непрямым» свободный билирубин называется потому, что не даёт прямой цветной реакции с диазореактивом, поскольку из-за плохой растворимости в воде он легко адсорбируется на белках плазмы крови.
Он быстро выводится из клеток Купфера и поступает в кровь, где адсорбируется альбумином. Такой комплекс называется свободным, непрямым, неконъюгированным билирубином. Он менее токсичен, нерастворим в воде и не проходит через почечный фильтр.
Этот комплекс с током крови поступает в печень, где распадается, а билирубин поступает в клетки печени – гепатоциты. Здесь билирубин взаимодействует с глюкуроновой кислотой с образованием глюкоронидбилирубина. В таком виде билирубин не токсичен, водорастворим и называется связанным билирубином (прямой, конъюгированный).
В клетках печени накапливается связанный билирубин, который поступает в желчный пузырь и входит в состав желчных пигментов. С желчью билирубин поступает в кишечник, где распадается. Освободившийся билирубин под воздействием ферментов микрофлоры толстого кишечника превращается в мезобилиноген. Небольшая часть мезобилиногена всасывается в кровь и по воротной вене доставляется в печень, где расщепляется до дипирролов, которые задерживаются печенью и не поступают в общий кровоток.
В сутки человек выделяет от 250 до 300 мг стеркобилиногена, который легко окисляется под действием света и воздуха до стеркобилина. Лишь небольшая часть билирубина (от 5 до 20 мг) выводится с калом в неизменном виде. Небольшая часть стеркобилиногена (до 5%) после всасывания через систему нижних геморроидальных вен попадает в большой круг кровообращения, минуя печень, и в таком виде выводится почками с мочой (около 4 мг в сутки).
Уробилиноген мочи и стеркобилиноген кала, окисляясь на воздухе, превращаются в уробилин и стеркобилин – пигменты мочи и кала.
Лабораторное определение уровня билирубина имеет большое значение в диагностике заболеваний печени, гемолитических состояниях (ГБН и др.).
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
В организме здорового человека с массой тела 70 кг каждый час разрушается около 1-2х108 эритроцитов, что, в перерасчете на массу гемоглобина, подвергающегося распаду, составляет, примерно, 6 г гемоглобина в сутки. Белковая часть гемоглобина разрушается до аминокислот, которые вновь используются клетками для синтеза белков. Катионы железа гема пополняют запасы железа в составе белка печени ферритина, а порфириновое кольцо гема разрушается до специальных продуктов – желчных пигментов (биливердин, билирубины и пигменты, образующиеся из билирубина в кишечнике).
Билирубин IXa
Билирубина диглюкуронид
Рис.19. Структура токсичного билирубина IXa и нетоксичного диглюкуронида билирубина.
Распад гемоглобина начинается, преимущественно, в клетках Купфера (печень) и клетках ретикулоэндотелиальной системы (РЭС) селезёнки (схема 3). Формирование промежуточных метаболитов распада гемоглобина (вердоглобин, биливердин, билирубин IXa) может происходить как в печени, так и в других органах, прежде всего в селезенке (схема 3), а конъюгация билирубина (образование диглюкуронида – только в гепатоцитах (схема 4)).
В качестве дополнительных источников билирубина выступают многочисленные гемопротеины, содержащиеся в значительных количествах во всех клетках тканей внутренних органов. Среди них особое место занимает белок скелетной мускулатуры и миокарда миоглобин, а также цитохромы.
Процесс превращения свободного (непрямого) билирубина, образующегося при разрушении эритроцитов и распаде гемоглобина в органах ретикулоэндотелиальной системы, в связанный, конъюгированный или прямой билирубин(билирубина диглюкуронид) в гепатоците (рис.20) осуществляется в три этапа (на рис.20 обозначены римскими цифрами):
I этап — захват билирубина (Б) печеночной клеткой после отщепления альбумина;
II этап — образование водорастворимого комплекса билирубин-диглюкуронида (Б-Г);
III этап— выделение образовавшегося связанного (прямого) билирубина (Б-Г) из печеночной клетки в желчные протоки.
Рис. 20. Функция гепатоцита в формировании и секреции прямого билирубина. Бн – свободный (непрямой) билирубин; Б-Г – билирубин-глюкуронид (связанный, или прямой билирубин); Мбг – мезобилиноген (уробилиноген).
Схема 3. Образование неконьюгированного (непрямого) билирубина.
Схема 4. Конъюгация билирубина в паренхиме печени.
В качестве источника глюкуроновой кислоты в этой реакции используется УДФ-глюкуроновая кислота:
Фермент УДФ-глюкуронилтрансфераза, подобно другим энзимам печени, подвергается индукции некоторыми ксенобиотиками. К таковым относятся в частности, широко используемые в клинической практике препараты барбитурового ряда (снотворные и седативные), некоторые противосудорожные препараты других фармакологических групп, транквилизаторы и пр.
В отличие от свободного билирубина, его конъюгированная форма хорошо растворима в воде. Конъюгаты билирубина, преимущественно в форме диглюкуронидов (до 75%), выделяются в желчь. При этом их транспорт направлен в сторону большей концентрации, т.е. против концентрационного градиента. Он обеспечивается специальной энергозависимой системой активного транспорта, встроенной в клеточную мембрану гепатоцитов. Дальнейший метаболизм билирубина связан с поступлением его в желчные пути и кишечник. В нижних отделах желчевыводящих путей и тонком кишечнике под воздействием микрофлоры происходит постепенное восстановление билирубина до мезобилиногена. Часть мезобилиногена. всасывается в кишечнике (уробилиноген) и по системе воротной вены вновь попадает в печень, где в норме происходит практически полное его разрушение до пирролов (схема 5).
Основное количество мезобилиногена из тонкой кишки поступает в толстую и здесь восстанавливается до стеркобилиногена при участии анаэробной микрофлоры (схема 5). Образовавшийся стеркобилиноген в нижних отделах толстой кишки (в основном в прямой кишке) окисляется до стеркобилина и выделяется с калом, определяя его окраску у здоровых людей. Лишь небольшая часть стеркобилиногена через геморроидальные вены попадает в систему нижней полой вены и в дальнейшем выводится с мочой (схема 5). Следовательно, в норме моча человека содержит лишь следы стеркобилиногена, не выявляющиеся клиническими лабораторными методами (за сутки его выделяется до 4 мг). К сожалению, до последнего времени в клинической практике стеркобилиноген, содержащийся в нормальной моче, продолжают называть уробилиногеном.
Схема 5. Превращение билирубина в кишечнике с образованием конечных продуктов в моче и кале.
При диагностике патологий пигментного обмена визуально оценивается интенсивность окраски мочи и кала больных, проводится количественное определение билирубинов (прямого, непрямого, общего) в плазме или сыворотке крови и уробилиногена в моче, иногда количественно определяется стеркобилин кала. При патологиях, сопровождающихся снижением концентрации эритроцитов (анемиях различной этиологии возникновения), обязятельным тестом является определение концентрации гемоглобина, и в качестве дополнительных, концентрация железа в крови человека.
Продолжительность
жизни эритроцитов составляет 120 дней, затем они разрушаются и освобождается
гемоглобин. Главными органами, в которых происходят разрушение эритроцитов и
распад гемоглобина, являются печень, селезенка и костный мозг, хотя в принципе
оба процесса могут происходить и в клетках других органов. Распад гемоглобина в
печени начинается с разрыва α-метиновой связи между I и
II кольцами порфиринового кольца. Этот процесс катализируется НАДФ-содержащей
ок-сидазой и приводит к образованию зеленого пигмента вердоглобина
(холеглобина):
В приведенных
структурных формулах здесь и далее в желчных пигментах М – метильная СН3-группа,
В – (—СН=СН2) – винильная группа и П – (—СН2—СН2—СООН)
– остаток пропионовой кислоты.
Как видно из
приведенных формул, в молекуле вердоглобина еще сохраняются атом железа и
белковый компонент. Имеются экспериментальные доказательства, что в этом
окислительном превращении гемоглобина принимают участие витамин С, ионы Fe2+и другие кофакторы. Дальнейший распад вердоглобина, вероятнее всего, происходит
спонтанно с освобождением железа, белка-глобина и образованием одного из
желчных пигментов – биливердина. Спонтанный распад сопровождается
перераспределением двойных связей и атомов водорода в пиррольных кольцах и
метиновых мостиках. Образовавшийся биливердин ферментативным путем
восстанавливается в печени в билирубин, являющийся основным желчным пигментом у
человека и плотоядных животных:
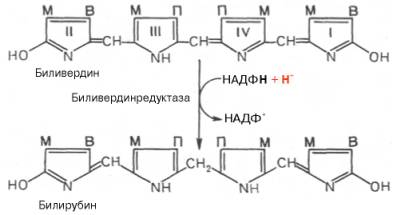
Основное
место образования билирубина – печень, селезенка и, по-видимому, эритроциты
(при распаде их иногда разрывается одна из метиновых связей в протопорфирине).
Образовавшийся во всех этих клетках билирубин поступает в печень, откуда вместе
с желчью попадает в желчный пузырь (см. главу 16). Билирубин, образовавшийся в
клетках системы макрофагов, называется свободным, или непрямым, билирубином,
поскольку вследствие плохой растворимости в воде он легко адсорбируется на
белках плазмы крови и для его определения в крови необходимо предварительное
осаждение белков спиртом. После этого билирубин вступает во взаимодействие с
диазореактивом Эрлиха.
В крови
взрослого здорового человека содержится относительно постоянное количество общего билирубина – от 4 до 26 мкмоль/л, в среднем 15 мкмоль/л.
Около 75% этого количества приходится на долю непрямого билирубина. Повышение
его концентрации в крови до 35 мкмоль/л приводит к желтухе. Более высокий
уровень билирубина в крови вызывает явления тяжелого отравления. Непрямой
билирубин, поступая с током крови в печень, подвергается обезвреживанию путем
связывания с глюку-роновой кислотой. В этом процессе принимают участие особый
фермент УДФ-глюкуронилтрансфераза и УДФ-глюкуроновая кислота, являющаяся
донором глюкуроновой кислоты. При этом к билирубину присоединяются 2 остатка
глюкуроновой кислоты с образованием сравнительно индифферентного комплекса –
билирубин-диглюкуронида, хорошо растворимого в воде и дающего прямую реакцию с
диазореактивом. В желчи всегда присутствует прямой билирубин. В крови
количество прямого и непрямого билирубина, а также соотношение между ними резко
меняются при поражениях печени, селезенки, костного мозга, болезнях крови и
т.д., поэтому определение содержания обеих форм билирубина в крови имеет
существенное значение при дифференциальной диагностике различных форм желтухи.
При желчнокаменной болезни в составе желчных камней наряду с основным их
компонентом – холестерином всегда обнаруживается непрямой билирубин. Вследствие
плохой растворимости в воде он выпадает в осадок в желчном пузыре в виде
билирубината кальция, участвующего в формировании камней.
Дальнейшая
судьба желчных пигментов, точнее билирубина, связана с их превращениями в
кишечнике под действием бактерий. Сначала глю-куроновая кислота отщепляется от
комплекса с билирубином и освободившийся билирубин подвергается восстановлению
в стеркобилиноген, который выводится из кишечника. В сутки человек выделяет
около 300 мг стеркобилиногена. Последний легко окисляется под действием света и
воздуха в стеркобилин. Механизм бактериальных превращений билирубина до
стеркобилина до конца еще не расшифрован. Имеются данные, что промежуточными
продуктами восстановления являются последовательно мезобилирубин и
мезобилиноген (уробилиноген). После всасывания небольшая часть мезобилиногена
поступает через воротную вену в печень, где подвергается разрушению с образованием
моно- и дипиррольных соединений. Кроме того, очень небольшая часть
стеркобилиногена после всасывания через систему геморроидальных вен попадает в
большой круг кровообращения, минуя печень, и в таком виде выводится с мочой.
Однако называть его уробилиногеном не совсем точно (см. главу 18). Суточное
содержание стеркобилиногена в моче составляет около 4 мг, и, пожалуй, именно
стеркобилиноген является нормальной органической составной частью мочи. Если с
мочой выделяется повышенное содержание уробилиногена (точнее, мезобилиногена),
то это является свидетельством недостаточности функции печени, например, при
печеночной или гемолитической желтухе, когда печень частично теряет способность
извлекать этот пигмент из крови воротной вены. Химически уробилиноген
(мезо-билиноген) неидентичен стеркобилиногену (уробилиногену) мочи.
Исчезновение стеркобилиногена (уробилиногена) из мочи при наличии билирубина и
биливердина является свидетельством полного прекращения поступления желчи в
кишечник. Такое состояние часто наблюдается при закупорке протока желчного
пузыря (желчнокаменная болезнь) или общего желчного протока (желчнокаменная
болезнь, раковые поражения поджелудочной железы и др.).
Таким
образом, количественный и качественный анализ желчных пигментов в моче может
представлять большой клинический интерес.
